-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
Research in Zoology
p-ISSN: 2325-002X e-ISSN: 2325-0038
2015; 5(1): 20-24
doi:10.5923/j.zoology.20150501.03

Body Shape Comparison of Kura Bleak (Alburnus filippii) in Aras and Ahar-Chai Rivers Using Geometric Morphometric Approach
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLPariya Jalili1, Soheil Eagderi1, Yazdan Keivany2
1Department of Fisheries, Faculty of Natural Resources, University of Tehran, Karaj, Iran
2Department of Natural Resources (Fisheries Division), Isfahan University of Technology, Isfahan, Iran
Correspondence to: Soheil Eagderi, Department of Fisheries, Faculty of Natural Resources, University of Tehran, Karaj, Iran.
| Email: |  |
Copyright © 2015 Scientific & Academic Publishing. All Rights Reserved.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

The body shape of two populations of Kura bleak (Alburnus filippii Kessler, 1877) from Aras and Ahar-Chai rivers was compared by geometric morphometric method. A total of 63 specimens of Kura bleak were collected from mainstream of Aras and its tributaries Ahar-Chai. Then, the lateral face of their left side were photographed and 13 landmark-points defined and digitized using TpsDig2 software. The obtained data were analyzed using multivariate analyses. The results revealed a significant difference in the body shape of the studied populations associating with body depth and the tip of the snout. The results also showed a higher morphological variation among the specimens of mainstream of Aras River compared to Ahar-Chai ones indicating availability of more diverse habitats. Thus, in a morphological comparative study of different populations of a single species in different habitats, the morphological variation of every population should be considered as a criterion and application of an exploratory multivariate analysis such PCA can help to better understand the phenotypic variations.
Keywords: TPS, Cyprinidae, Caspian Sea Basin, Iran
Cite this paper: Pariya Jalili, Soheil Eagderi, Yazdan Keivany, Body Shape Comparison of Kura Bleak (Alburnus filippii) in Aras and Ahar-Chai Rivers Using Geometric Morphometric Approach, Research in Zoology , Vol. 5 No. 1, 2015, pp. 20-24. doi: 10.5923/j.zoology.20150501.03.
Article Outline
1. Introduction
- The differences between populations of a single species can indicate differences in habitat and behavioral characteristics, because the aquatic organisms, e.g., fishes require to adapt to their environmental conditions for better functioning of their biological systems [1]. Therefore, morphological adaptations to environmental conditions along with geographical isolation can provide a crucial information on the evolutionary trend of this organisms particularly in aquatic ecosystems such as rivers [2]. River ecosystems includes only 0.001% of water resources of our planet, nevertheless includes almost half of the fish species i.e. its species diversity is 7500 times greater than other aquatic systems [3]. Two main factors for such a diversity are geographical isolation and diversity of the environmental conditions [3, 4].Rivers have a high variation in terms of the hydrological and biological features, which can effect the biological characteristics of the aquatic organisms. Hence, due to differences in habitat characteristics of a river ecosystem and the possibility of morphological diversity due to variation of the habitat conditions, this study was aimed to compare the body shape of two populations of Kura bleak, Alburnus filippii Kessler, 1877, (Fig. 1A) inhabiting the mainstream and a tributary of Aras River to answer this question that whether morphological features of the tributary population is different from that of the mainstream? Two populations of fish with a common origin may show a different local adaptations due to environmental factors [5]. Therefore, the results of this study can provide an important information on the response and the evolutionary trend of this species. The population living in the mainstream of Aras River (at Khomarlu; 39°11'37"N and 47°02'03"E) with a width of 30 m, a depth of 1.5-2 m and a velocity of 1.56 m/s and a second population from Ahar-Chai River (a tributary of Aras River) (38°27'02"N and 47°02'12"E) with a width of 4 m, a depth of 15-30 cm and a velocity of about 1 m/s were selected. In addition, there is a high geographical distance between these two populations.
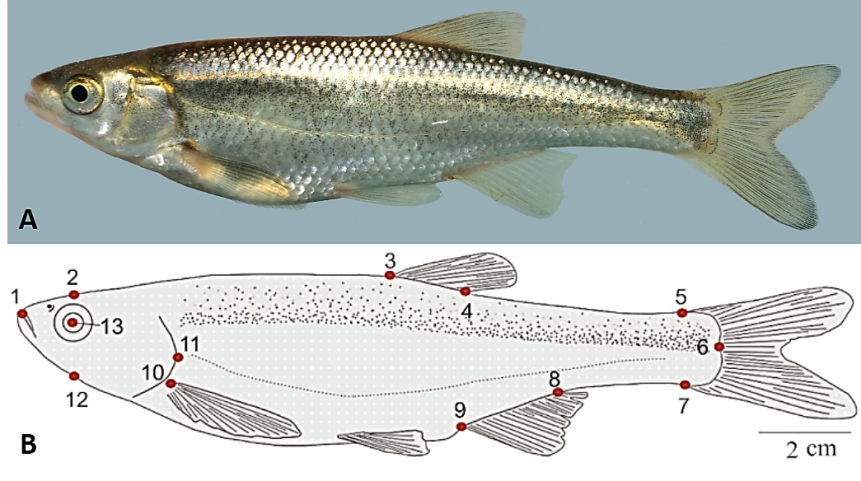 | Figure 1. (A) Alburnus filippii from Aras River and (B) defined landmark points to extract the body shape data: (1) anterior-most point of the snout tip on the upper jaw, (2) dorsal edge of the head perpendicular to the center of eye, (3) anterior and (4) posterior end of the dorsal fin base, (5) posterodorsal end of caudal peduncle at its connection to caudal fin, (6) posterior most of body lateral line, (7) posteroventral end of caudal peduncle at its connection to caudal fin, (8) posterior and (9) anterior ends of the anal fin base, (10) dorsal origin of pectoral fin, (11) posterior edge of operculum, (12) ventral edge of the head perpendicular to the center of eye and (13) center of eye |
2. Material and Methods
- A total of 63 specimens of Kura bleak (A. filippii) including 32 specimens from main canal of Aras River and 31 specimens from Ahar-Chai River were collected in June 2013 by electrofishing (Fig. 2). The specimens were anesthetized in MS-222 and then the specimens of almost similar sizes (with standard length more than 90 cm) were selected to remove the complications of body shape variation due to allometric growth patterns and also the selected specimens had no deformity in terms of body shape. The left sides of fresh collected specimens were photographed using a copy-stand equipped with a digital camera (Kodak EasySharw Z650 with 6 MP resolution).
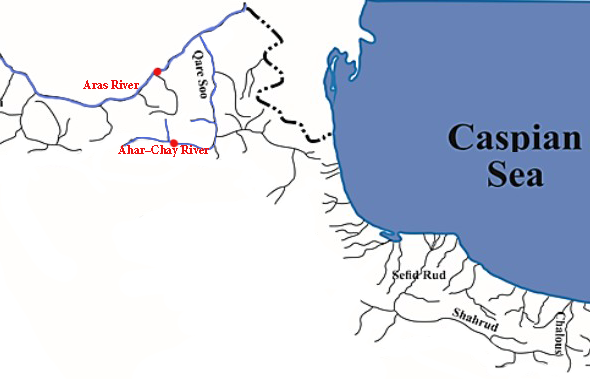 | Figure 2. The studied locality of Alburnus filippii populations in Aras and Ahar-Chai rivers |
3. Results
- Principal components analysis was explained 88% of shape variations by the first four PC axes extracted from the variance-covariance matrix (PC1 = 58.5%, PC2 = 14.05%, PC3 = 8.85% and PC4 = 6.77%). The graph of the first two components with a total variance of 72% revealed that the mainstream population of Kura bleak has a higher morphological variation than that of the Ahar-Chai River (Fig. 3). Distribution of the specimens in PCA graph showed the presence of two morphological types in the mainstream of the Aras River, which first type along with the specimens of the Ahar-Chai population were located at negative part of the PC2 axes showing a similar body pattern. The specimens of this morph have a deeper body and a longer caudal peduncle. The second morph type of Kura bleak belonged to the specimens of the mainstream population, was positioned in the positive part of the PC2 axes. The members of this morph type have a lower body depth, a shorter caudal peduncle, a pectoral fin with a ventral position and snout and eye with a dorsal position (Fig. 3).
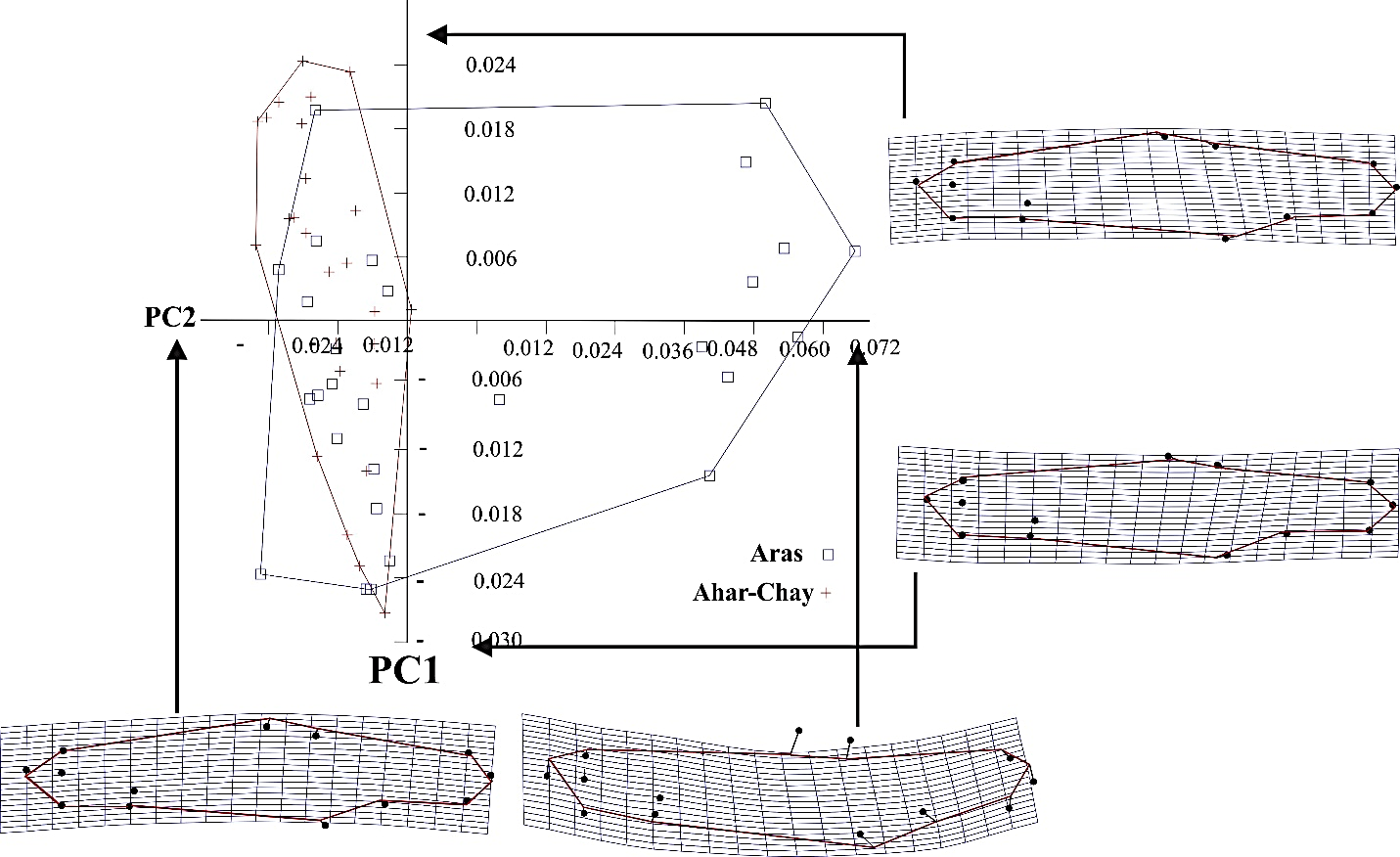 | Figure 3. Scatter plot of individual scores of the first two principal components of the studied populations of Alburnus filippi (deformation grids show the most extreme positive and negative shape deformations of Kura bleak) |
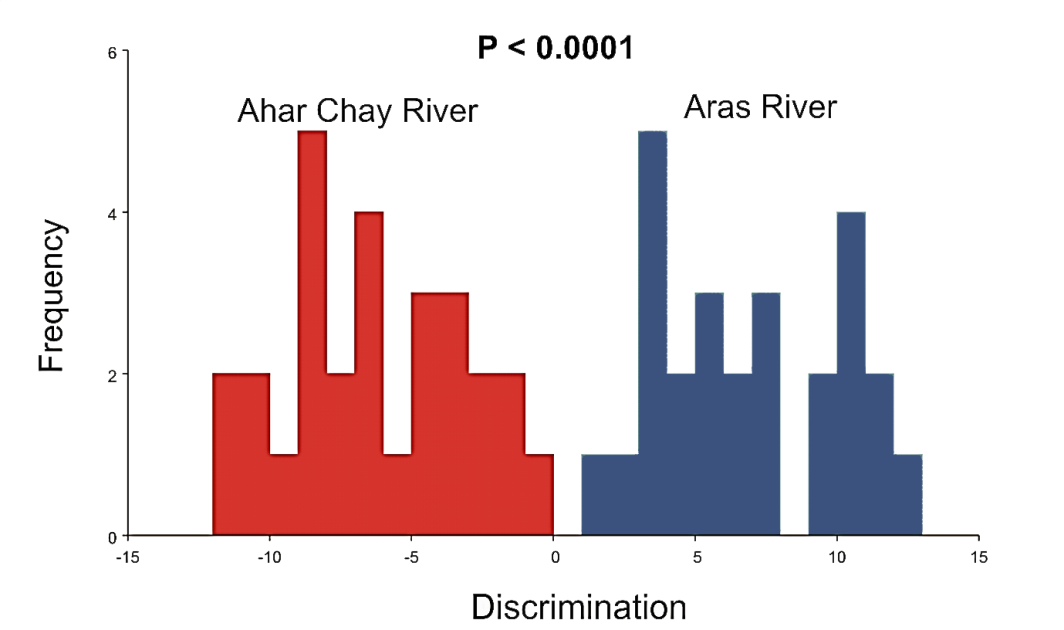 | Figure 4. Graphs of the discriminant function analysis of the two populations of Alburnus filippi |
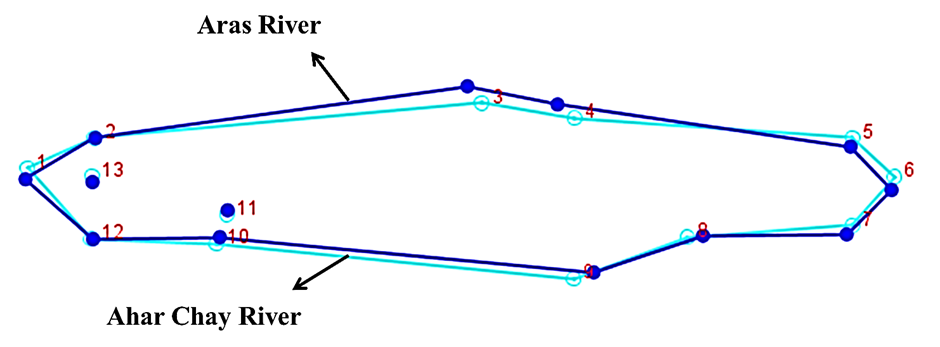 | Figure 5. Wireframe graph showing the consensus body shape differences of two populations of Alburnus filippi |
4. Discussion
- The results of the present study showed significant morphological differences between populations of Kura bleak in the mainstream of Aras River and in its tributary, Ahar-Chai River, indicating the presence of a higher morphological diversity in the mainstream. Different morphological characteristics of a single species populations can be due to either genetic differentiation or phenotypic plasticity in response to environmental parameters of their habitat [13-16]. Since the habitat factors have a significant effect on morphology of fish species [17, 18], therefore, the observed morphological differences in the mainstream of Aras River, with the similar origin, can be related to availability of more diverse habitats, i.e., in the mainstream more niches are available for fish and consequently more variation in body shape can be observed. Environmental factors through natural selection can increase the efficiency of a phenotype among the members of a population and thus is led to morphological isolation in different habitats [2, 19].Phenotypic variations among fishes in various habitat may be viewed as important only if any differences in morphology translate into an increased in fitness among the different groups [4]. Body shape differences between two habitats often reflect variations in the swimming and feeding of fishes [20, 27, 28]. Fishes living in an environment with strong water current will have to conform to this habit. The lower body depth of second morphological type of the mainstream of the Aras River would then suggest a more active habit, i.e., a habitat with strong water current. An elongated body shape with lower body depth, i.e., a fusiform shape reduces drag [20] and hence reduces the energy expenditure necessary to maintain position in flowing water [21, 22]. Also, in the second morphological type of the mainstream population, the mouth position was relatively upturned. Intraspecific trophic diversification is well-known in fish species [23-25]. Therefore, the observed differences in mouth position among studied specimens could reflect differences in feeding [20].
5. Conclusions
- Based on the results of the present study, we can answer this question; whether morphological features of the tributary population is different from that of the mainstream?. A significant morphological differences between populations of the mainstream of Aras River and its tributary, Ahar-Chai River was observed. The results revealed two morphological types in mainstream that one of them has similar morphospace with that of the tributary and this could be due to availability of more diverse habitats in the main-stream and consequently more variation in body shape. [2, 19].In addition, our findings suggest that in a morphological comparative study of different populations of a single species in different habitats, the morphological variations of populations should be considered in relation to inter-population comparison. Hence, application of an exploratory multivariate analysis such as principal component analysis (PCA), to find out the patterns in a dataset, can help to better understanding of the phenotypic variation among organisms instead of comparative analysis [8, 26]. As shown in this study, DFA analysis revealed a significant differences between the two populations, but PCA revealed that the two distinct morphological types are found among the members of the mainstream of the Aras River population that one of them is similar to that of its tributary.
ACKNOWLEDGEMENTS
- We would like to thank all the students who helped in fish collection and field work. This study was financially supported by the University of Tehran and Isfahan University of Technology.