-
Paper Information
- Previous Paper
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Plant Research
p-ISSN: 2163-2596 e-ISSN: 2163-260X
2012; 2(2): 14-21
doi: 10.5923/j.plant.20120202.03
Evaluation of Early and Late/Intermediate Maize Varieties for Grain Yield Potential and Adaptation to a Southern Guinea Savanna Agro-ecology of Nigeria
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-Text HTML
Full-Text HTMLBello O. B. 1, Abdulmaliq S. Y. 2, Ige S. A. 3, Mahamood J. 4, Oluleye F. 5, Azeez M. A. 6, Afolabi M. S. 7
1Department of Biological Sciences, Fountain University, Osogbo, Osun State, Nigeria
2Department of Agronomy, Ibrahim Badamasi Babangida University, Lapai, Niger State, Nigeria
3Department of Agronomy, University of Ilorin, Ilorin, Nigeria
4Lower Niger River Basin Development Authority, Ilorin, Kwara State, Nigeria
5Kwara State University, Malete, Ilorin, Kwara State, Nigeria
6Department of Plant Biology, Ladoke Akintola University of Technology, Ogbomoso, Nigeria
7Department of Crop Science, Landmark University, Omuaran, Kwara State, Nigeria
Correspondence to: Bello O. B. , Department of Biological Sciences, Fountain University, Osogbo, Osun State, Nigeria.
| Email: |  |
Copyright © 2012 Scientific & Academic Publishing. All Rights Reserved.
Drought-tolerant maize varieties belonging to two different maturity (10 early and 10 late/intermediate) groups were evaluated for yield and other related characters in the southern Guinea savanna (SGS) of Nigeria for two years (2009 and 2010). The differences among genotypes between and within maturity groups differed significantly (P<0.01) for grain yield, plant height, days to anthesis and silk. The effect of year x maturity group and year x maturity within group interactions was highly significant (P<0.01) only for grain yield. The rainfall patterns were favourable in both cropping years with comparable values of growth parameters. Late/intermediate maturing varieties (TZL COMP1-W C6 F2, SUWAN-1-SR-SYN, TZB-SR, OBA SUPER I, EV 8435-SR) out-yielded early maturing ones with yield advantage of 34.29% and taller by 17.04% compared to early ones. However, early varieties were early to anthesis with 6.57% advantage over late/intermediate cultivars. Four early varieties (DMR-ESR Y CIF2, AC 90 POOL 16 DT, STR, TZE-W DT STR C4 and ACR 95TZE COMP4 C3) were superior for grain yield withn a range of 4.39 to 4.68 t/ha. These genotypes could be used either as cultivar per se to escape the prolonged moisture stress during the later part of the cropping season or introgressed with favourable cultivars for high yield adaptable to drought-prone areas in the southern Guinea savanna agro-ecology.
Keywords: Maize yield, early varieties, late/intermediate varieties, maturity group
1. Introduction
- Maize (Zea mays L.) is an important staple food crop and provides bulk of raw materials for the livestock and many agro-allied industries in the world (Bello et al., 2010; Randjelovic et al., 2011).). The savanna agro-ecology of Nigeria has a great potential for food production because of its high solar radiation that favours maize production. In the southern guinea savannah (SGS) however, maize is grown twice due to bimodal rainfall pattern (a short early growing season followed by fairly long late season). Early maize varieties are usually planted at the onset of rainy season before it’s fully established (March/April), and matured sooner than the traditional crops (sorghum [Sorghum bicolor (L.) Moench] and millet [Pennisetum glaucum (L.) R. Br.]). This succors in filling the hunger gap in July when all food reserves have been depleted after the long dry period in the zone. On the other hand, the late season crop is planted during the second cycle of rains (July/August). While the short early season is usually characterized by abrupt cessation of rains during crop cycle, the late season is normally affected by terminal drought. The occurrence of extreme environmental events impose different degrees of drought stresses on crops thereby affecting growth duration, plant size, dry matter accumulations, assimilate reserves and partitioning to grains. As the episodes of water resources for agronomic uses become more limiting, the development of drought-tolerant (DT) lines becomes increasingly necessary. Drought occurring during or shortly before flowering in crops, the estimated yield loss may be in the range of 21 to 50% (Olaoye et al., 2009). The soil in the SGS is also fragile with low organic matter, poor buffering and water holding capacity, resulting in low nitrogen availability (Fakorede et al., 2001; 2003). Since the timing of mid-season drought is unpredictable, early maize cultivars that can tolerate the effects of reduced moisture supply during flowering could reduce farmers’ risk in drought-affected ecologies (Olaoye et al., 2009; Hussain et al., 2011). Early maturing DT maize varieties are ideal for intercropping by providing less competition for moisture, light, and nutrients than late maturing ones. They also offer flexibility in planting dates, which enables: (i) multiple plantings in a season to spread risk of losing a single crop to drought (ii) late plantings during delayed onset of rainfall, and (iii) avoidance of known terminal drought periods during the cropping season (CIMMYT, 2000). occurring The occurrence of drought in most parts of West and Central Africa, has now made the production and utilization potential of DT maize varieties attract the attention of the national and international researchers to developi, testing and transferr high yielding and adapted maize cultivars to farmers (Badu- Apraku et al., 2003; Olaoye and Omueti 2006).The release of improved drought-tolerant maize varieties by these research institutes has created assurance for increased maize productivity in the savanna agro-ecologies. These could not only be achieved by promoting rate of adoption of improved maize cultivars by the farmers, but also provide farmers opportunities to overcome the challenges to maize production, thereby improving food security in the West and Central Africa. From 1987 to date, the regional early and late/intermediate varietal trials have been organized by West and Central Africa Maize Collaborative Research Network (WECAMAN) and the Maize and Wheat Improvement Center (CIMMYT), Mexico as a vehicle for systematic testing and dissemination of elite maize across West African sub-region. They were on germplasm exchange among maize scientists in the sub-Sahara of West Africa with the aim of offering the opportunity to identify varieties suitable to their peculiar growing condition. Evaluation of extra early (80-85 days), early (90-95 days) and late/intermediate (100-105 days) maturing groups have formed part of their varietal trials in the marginal environments of the region under the auspices of International Institute of Tropical Agriculture (IITA) Ibadan, Nigeria (Badu- Apraku et al., 2003; Sallah et al., 2004; Olaoye and Omueti, 2006; Oluwaranti et al., 2008). Availability of the early and extra-early varieties is a strategy for adapting maize to the gradually shortening rainy season, so that maize could tolerate or/and escape the drought stress that occurs during the grain-filling stage in the late season. These cultivars are endowed with favourable genes for high yield (about 20- 50% yield increase than other maize varieties) potential and stability across a broad range of water availability (Kucharik, 2008; Olaoye et al., 2009). On this evidence, assessment of early maturing DT varieties that fully explored the potential growing season and fit into the erratic rainfall pattern in the ecology is precondition to increasing maize yields. The present study was therefore conducted to evaluate the performance of early and late/intermediate DT maize maturity groups, with the view to identifying cultivars that are high yielding and adapted to marginal and drought-prone ecologies for farmers’ use.
2. Materials and Methods
- Collection of planting materialsThe genetic materials used for this study was a set of drought-tolerant maize varieties belonging to two different maturity (10 early and 10 late/intermediate) groups. These varieties were developed by West and Central Africa Maize Collaborative Research Network (WECAMAN) which was originally part of the international trials coordinated by the IITA Ibadan, Nigeria. The popular local variety, AFO and Composite-Y were used as checks for both early and late/intermediate maturity groups respectively. The genotypes were evaluated for two years (2009 and 2010) during late growing seasons at the Lower Niger River Basin Development Authority station, Oke-Oyi, Ilorin, Nigeria (Latitude 8° 30’N, 8°36’E and Longitude 4° 31’N, 4° 33’E). Planting was carried out on 21st July, 2009 and 27th July, 2010 respectively.Soil analyses and rainfall dataThe physico-chemical characteristics of the soil at the experimental site were determined by collecting soil samples randomly at the depths of 0-15cm and 15-30cm respectively with an auger, and analyzed in the laboratory before planting in both cropping seasons (Table 1). The collected samples were air-dried and passed through 2mm sieve to remove large particles, debris and stones. The sieved samples were analyzed for pH in 1:1 soil to water ratio using the Coleman’s pH meter. Organic carbon was determined by Walkley and Black procedure (Nelson and Somers, 1992). Total nitrogen was determined by the micro Kjeldahl method (Bremner, 1965), while available phosphorus was extracted by Bray’s P1 method (Bray and Kurtz, 1945) and read from the atomic absorption spectrometer. Exchangeable Ca, Mg, K, Na and effective cation exchangeable capacity (ECEC) were analyzed using Atomic Absorption Spectrophotometery (IITA, 1989), while textural analysis was by hydrometer method. Rainfall distribution data for 2009 and 2010 were also recorded (Figure 1).
|
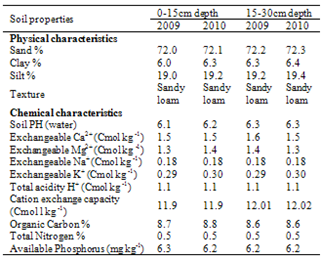
 | Figure 1. Monthly rainfall distribution pattern for Ilorin in 2009 and 2010 |
3. Results
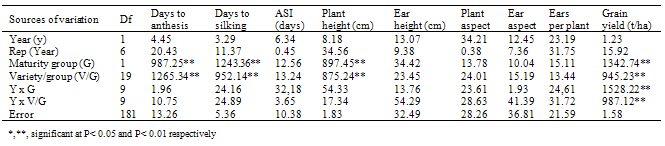
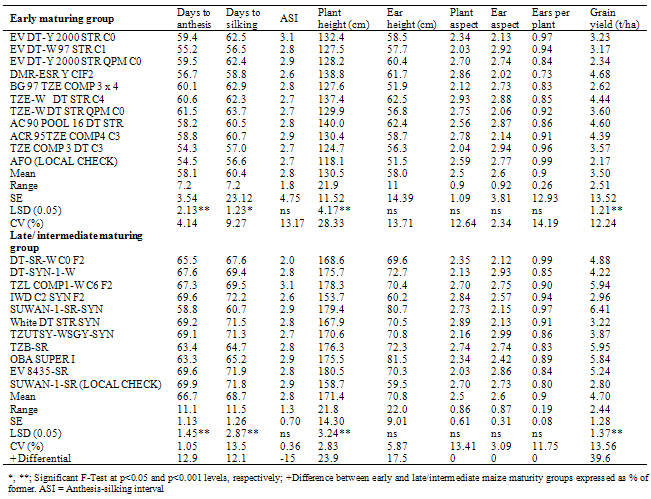
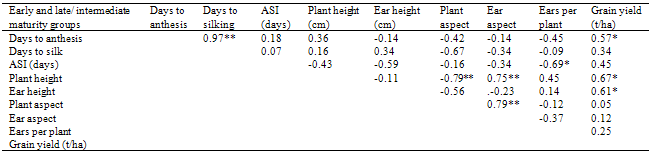
- Distribution of rainfall and soil analysisIn the cropping seasons of 2009 and 2010, there were false start of rain from February to March and a break in July, 2010. This then followed by adequate and evenly distribution throughout the flowering/grain filling periods of July to October. In each of the growing year, there was a significant rainfall drop (August break) in August, 2010 and October 2009. (Figure 1). However, the physico-chemical analysis of 0-15 and 15-30cm soil depth showed that the soil at the experimental area was comparable in both depths in both years (Table 1). The particle size analysis of the soil type was sandy loam with a high proportion of sand (72.2%) but less clay (6.3%), silt (19.2%). On the chemical characteristics, the soil was slightly acidic (pH of 6.2) with adequate cation exchange capacity (11.90 Cmol kg-1). Although the total nitrogen (0.5%), organic carbon (8.7%), calcium (1.5 Cmol kg-1) and magnesium (1.3 Cmol kg-1) values were low, there was a high concentration of available phosphorus (6.2 Cmol kg-1). Mean squares from combined analysis of varianceThe differences among genotypes between and within maturity groups differed significantly (P<0.01) for grain yield, plant height, days to anthesis and silk (Table 2). The effect of year x maturity group and year x maturity within group interactions were highly significant (P<0.01) only for grain yield. The sources of variation however were non- significant for all the characters in both years. The rainfall patterns were favourable in both cropping years (Figure 1) with comparable values of growth parameters. Genotypic performance for yield and other agronomic charactersRanges in the means for grain yield, ears per plant, anthesis-silking interval (ASI), plant and ear aspect scores were lower in the late/intermediate maturity genotypes than early varieties (Table 3). While ranges in the means for ear height, days to anthesis and silk were high, reverse was the case for the values of plant height with comparable results. Although, early varieties were early to anthesis with 6.57% advantage over late/intermediate cultivars, late/intermediate varieties had grain yield advantage of 34.29% and taller by 17.04% compared to early varieties regardless of the growing year. Among early varietal group, DMR-ESR Y CIF2 was superior with highest grain yield followed by AC 90 POOL 16 DT STR, TZE-W DT STR C4 and ACR 95TZE COMP4 C3 in that order. However, most of the late/intermediate maturing varieties out-yielded early maturing ones. In late/intermediate maturing group, TZL COMP1-W C6 F2, SUWAN-1-SR-SYN, TZB-SR, OBA SUPER I, EV 8435- SR yielded higher with yield advantage of 29.06% compared with the best (DMR-ESR Y CIF2) in the early group. It is noteworthy that the two local varieties as checks (SUWAN-1-SR and AFO) were the poorest for grain yield. On the other hand, similar and moderate ASI, ears per plant as well as plant and ear aspects scores were recorded between and within maturity groups. Among early maturity group, ranking based on grain yield and days to silk showed that DMR-ESR Y CIF2 ranked top followed by AC 90 POOL 16 DT STR (Table 4). Consequently, ACR 95TZE COMP4 C3 and Syn E2 that had top entry for grain yield, ranked a distant 10th because of lateness in anthesis. Check variety (AFO) that was the poorest for grain yield had rank entry of 6th based on its earliness to anthesis. However, among late/intermediate maturity groups, SUWAN-1-SR-SYN ranked top followed by TZB-SR., TZL COMP1-W C6 F2 and OBA SUPER I in that order (Table 5). Check variety, SUWAN-1-SR that was the poorest for grain yield also ranked a distant 10th based on its lateness to anthesis. It is important to note that the local variety, SUWAN-1-SR that was the lowest grain yielding had yield advantage of 129% after being improved to form SUWAN-1-SR-SYN.
|
|
|

|
4. Discussion
- Precipitation pattern of rainfall has great impact in the expression of plants’ potentials during period of flowering/ grain filling of the crop growth cycle, especially maize. It also plays significant role in stimulating plants to speed up the process of maturation. Rainfall distribution and amount was probably the single most important environmental factor that affected overall crop performance in this study. The rainfall patterns were favourable in both cropping years which resulted to comparable values of the agronomic parameters (Figure. 1). For example, soil moisture that was not limiting at anthesis during the two growing years created favourable environments for seed set and translocation of assimilates to grain filling. Though, soil analysis of the experimental site was sandy loam and slightly acidic favourable for maize growth, the soil nutrients especially nitrogen and organic matter were low. This may be due to continuous cropping of the land over four years without fallowing. With the appreciable application of fertilizer, appropriate weeding and favourable rainfall probably gave individual genotype ample opportunity to express their yield potentials. On the other hand, the differences in performance of the genotypes both between and within each maturity groups for grain yield, plant height, days to anthesis and silking confirmed the diversity of the genotypes and their differences for these characters. Year x maturity and year x maturity within group interactions that were significant only for grain yield underscored the differences in genetic makeup of the genotypes which have pre-requisite advantages in breeding for improved grain yield. Furthermore, this wide variability observed for these yield parameters showed that they were quantitatively inherited and offered way for further improvement through selection (Bello and Olaoye, 2009). These results were expected as the varieties were of different maturity classes (early and late/intermediate maturity), different genetic backgrounds and bred for different traits such as drought tolerance, downy mildew resistance and streak resistance (Oluwaranti et al., 2008).Plant height is not only important for breeding of new varieties of maize, for green and dry matter production, but also for grain yield. The present result revealed highly significant variability in both early and late/intermediate varieties for plant height (Table 2). Plant height has been observed to be controlled by the expression of many genes and the interactions between these genes (Yamakawa et al., 2006; Tahir et al., 2008). Many researchers have shown highly significant variability in plant height in various maize genotypes (Salami et al., 2007; Naushad et al., 2007; Nazir et al., 2010; Iqbal et al., 2010a; Iqbal et al., 2010b). High correlation of grain yield with plant height was earlier reported by other researchers (Bello et al., 2010; Nazir et al., 2010). Nazir et al., (2010) also reported that plant height was positively correlated with days to flowering morphologically, as internodes’ formation stops at floral initiation, and that early flowering maize varieties are usually shorter in height.Ear height has also been described to be one of the most important selection criteria in most breeding programmes especially the root and stock lodging (Esechie et al., 2004).. High ear position could be susceptible to root and stock lodging, therefore the plant breeders usually prefer selecting for lower ear position in maize (Esechie et al., 2004; Salami et al., 2007). Early varieties were shorter compared to late/intermediate with high ear heights in this study. Some early varieties (DMR-ESR Y CIF2, AC 90 POOL 16 DT STR, TZE-W DT STR C4 and ACR 95TZE COMP4 C3) that had short ear height were high yielding. Previous researcher reported reduced ear height and increased grain yield in maize (Olakojo and Olaoye, 2005; Salami et al., 2007; Nazir et al., 2010). Lower plant and ear height augments plant lodging resistance in maize with increase grain yield (Esechie et al., 2004). While high vertical root-pulling resistance (lodging resistance) took up more N and utilized it more efficiently, better agronomic performance and higher yield resulted (Kamara et al., 2003; Liu and Wiatrak, 2011). Therefore, these early varieties could also be N-efficient or tolerant to N deficiency. Consequently, low ear heights could be suited to the conditions of small-holder farmers because the farmers hire small children and women for harvest. Harvesting using children becomes less cumbersome, and losses due to “gleaning” are reduced. Accessibility of short and early maturing varieties to small-holder farmers has a good advantage, because higher yields could result from small cultivated area with increase in plant density (Nik et al., 2011). Furthermore, shorter plants attached with less and short leaves require high plant density, and could compete for light and soil nutrients for high yield favorably than the taller ones. For large-scale commercial farmers utilizing combined harvester, the operation is also less tedious. With the lodging resistance, large-scale farmer does not need to recruit extra hands to pick ears from lodged plants. (Ahmad et al., 2011). also reported that availability of earlier maize varieties with shorter plant height, lower leaf number, upright leaves, smaller tassels and better synchrony between male and female flowering time, has enhanced the ability of maize to face high plant populations without showing excessive barrenness. Consistent number of days (3 days) was recorded for anthesis-silking interval (ASI) among early and late/ intermediate maturity groups in both years. This indicated an interval of 3 days between pollen shed and silk intrusion in the genotypes. Bello and Olaoye (2009) described ASI as a measure of nicking (synchronization) of pollen shed with silking. ASI has been reported to be a valuable diagnostic trait for cultivar performance under stress than days to silking per se, since it is largely independent of maturity differences among cultivars (Ibikunle et al., 2009; Abdalla et al., 2010). The number of ears per plant decreased significantly as ASI increased, and this trait was a major factor that contributed to differences between the top and lowest yielding genotypes under drought stress ((Bello and Olaoye, 2009). Genotypes with reduced ASI will allow for better fertilization and good cob fill (Ibikunle et al., 2009; Ahmad 2011) Therefore, selection for reduced ASI in tropical maize varieties has been shown to be correlated with improved yields under drought stress (Ibikunle et al., 2009; Abdalla et al., 2010; Ahmad 2011). Plant and ear aspects are also vital in determining varietal acceptability under farmer’s condition. Our result showed that among early and late/intermediate maturity groups, plant and ear aspects were fair in overall phenotypic appeal (2.6), but not significantly different in both years (Table 3). This indicated that these genotypes were not greatly affected with prevalent diseases during the two cropping seasons.Comparison between the early and late/intermediate maturity showed that most of the late/intermediate varieties out yielded early varieties. Late/intermediate varieties were late to maturity, higher in plant and ear heights and yield, compared to early group. It is generally recognized that longer maturity variety produced greater yield to enable for a long duration in metabolic transformation into grain and stover yields (Agele, 2006; Golbashy et al., 2010; Song et al., 2010; Hussain et al., 2011; Wang et al., 2011). Early maturing varieties on the other hand, required fewer corn heat units to reach flowering, while late maturing cultivars exhibited extended vegetative period. Therefore, early flowering maize plants are smaller and have fewer leaves with low grain yield compared with late cultivars (Shi et al., 2008; Akbar et al. 2009; Kamara et al., 2009; Wang et al., 2010; Khan et al., 2011). However, grain yield and earliness to silk are two important characters that can be used in ranking genotypes for their suitability as cultivars especially in a drought–prone ecology such as savannas (Table 4). Earliness to anthesis and/ or silking allows short growth duration and maturity; and these could constitute important attributes of drought escape which often make earlier maturing maize genotypes adapt better to late season moisture stress than late maturing ones (Olakojo and Olaoye, 2005; Naushad et al., 2007; Salami et al., 2007; Akbar et al., 2008; Shi et al., 2008; Nazir et al., 2010; Majid et al., 2010). This therefore showed a direct relationship between grain yield and maturity irrespective of the group. Reports from WECAMAN regional trials (Anon, 2001) also indicated that this situation is not peculiar to this study. Therefore, four early varieties (DMR-ESR Y CIF2, AC 90 POOL 16 DT STR, TZE-W DT STR C4 and ACR 95TZE COMP4 C3) that combined low plant and ear heights and earliness to anthesis with grain yield were likely related to their earliness in reaching maturity which permitted the genotypes to better utilize the available soil moisture during anthesis (Olaoye et al., 2009). They probably contain certain attributes of drought escape and could adapt better to late season moisture stress than late maturing counterparts especially in the erratic rainfall pattern of the SGS agro-ecology (Shi et al., 2008; Wang et al., 2011). Nonetheless, AFO that was earliest to anthesis (54.5 days) among all the genotypes could be hybridized with higher yielding varieties for yield improvement in the sub-region. In conclusion, late/intermediate varieties (SUWAN-1-SR-SYN, TZB-SR., TZL COMP1-W C6 F2 and OBA SUPER I) which combined grain yield with earliness to silking and appreciable ASI and ear aspect among others could be grown in the zone. These superior varieties that flowered and matured earlier with high yield could be used to escape the prolonged moisture stress during the later part of the cropping season. They could be tested on the farmers’ field for adaptation and adoption by the farmers. They could also serve as potential sources of unique combinations of favorable alleles for developing high yielding varieties adapted to drought affected areas in West and Central Africa.