-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
Marine Science
p-ISSN: 2163-2421 e-ISSN: 2163-243X
2014; 4(2): 49-57
doi:10.5923/j.ms.20140402.04
Paleoecology of Upper Cretaceous Sediments in Central Iran, Kerman (Bondar-e Bido Section) Based on Ostracods
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLBita Ghasem Shirazi1, Lida Bakhshandeh2, Abdollah Yazdi1
1Department of Geology, Islamic Azad University, Kahnooj Branch, Kerman, Iran
2Department of Geology, Science and Research Branch, Islamic Azad University, Tehran, Iran
Correspondence to: Bita Ghasem Shirazi, Department of Geology, Islamic Azad University, Kahnooj Branch, Kerman, Iran.
| Email: |  |
Copyright © 2014 Scientific & Academic Publishing. All Rights Reserved.
In order to assess the biological potential of ostracods in response to environmental processes, in this study, the Upper Cretaceous sediments of Bondare Bido Section, has been studied. The study Section is located at 45 km away in South-West direction of the Kerman (Iran). The area between northern latitude 57˚17' 48 and western longitude 29˚49'44. Due to high sensitivity to environmental changes, the Ostracods are reliable indicators to determine water depth in Upper Cretaceous. On the bases of identified microfossils, the Upper Cretaceous sediments of the study area are Turonian-Maastrichtian in age. These deposits have been biostratigraphically and paleoecologicall point of view evaluated and boundary between stages are determined. Here we report 13 genuses of well preserved ostracods belong to two family Podocopida and Platycopida. The assemblage of ostracods and foraminifera are good indicator of ecological environment during deposition of Cretaceous sediments. The abundance of genus and species of planktonic foraminifera and the presence of some benthic foraminifera and ostracods, may indicate relatively deep and open marine (hemiplegic) environment for these deposits.
Keywords: Paleoecology, Central Iran, Ostracod, Upper Cretaceous, Bondare Bido Section
Cite this paper: Bita Ghasem Shirazi, Lida Bakhshandeh, Abdollah Yazdi, Paleoecology of Upper Cretaceous Sediments in Central Iran, Kerman (Bondar-e Bido Section) Based on Ostracods, Marine Science, Vol. 4 No. 2, 2014, pp. 49-57. doi: 10.5923/j.ms.20140402.04.
Article Outline
1. Introduction
- It is for the first time that foraminifera & ostracods from the Bondare Bido Section were studied by the authors. The Cretaceous sediments have covered vast area of Kerman. These deposits outcropping at north, north- west and south of Kerman city as an important mountains of Reef limestone. Samples have been systematically taken from Bondare Bido section and subjected for investigation.Most of the ostracods have been identified at genus level and for determination of their age we used present foraminifera, associated with ostracods. The identified index fossils of foraminifera include:Whiteinella archaeocretacea Pessagno, Marginotruncana sigali (Reichel), Dicarinella concavata (Brotzen), Dicarinella asymetrica (Sigal), Muricohedbergella holmdelensis Olsson, Contusotruncana fornicata (Plummer), Macroglobigerinelloides bollii (Pessagno), Globotruncana ventricosa, Globotruncanita elevata (Brotzen), Radotruncana calcarata (Cushman), Globotruncanella havanensis (Voorwijk), Globotruncana aegyptica Nakkady, Gansserina gansseri (Bolli), Contusotruncana contuse.The assemblage of index foraminifera fossils suggest Turonian-Maastrichtian age for Bondare Bido Sections.
2. Methodology
- The ostracods were studied in three stages including library, field and laboratory studies. In field study, a section was chosen and sampling systematically was done. In the present study, 58 samples of Upper Cretaceous sediments with a thickness of 265m has have been studied. About 300g of each sample was soaked in water for a few days, and then samples 12cc of 15% hydrogen peroxide solution then samples were macerated using, sieves with a diameter of 30 and 60 meshes.The dried samples were poured into a tray and ostracods were picked up under optical microscope. After studying and Scanning Electron Microscopy was applied for identification.
2.1. The Geographical Location and Accessibility of Bondare Bido Section
- The Bondare Bido Section is located at 45 km away in South-West direction of the Kerman. The area between northern latitude 57˚17' 48" and " western longitude29˚49'44, this section is located in 1:100,000, geological map of Rayen.The study area is accessible through the asphalt road of Kerman-Bam (Figure 1).
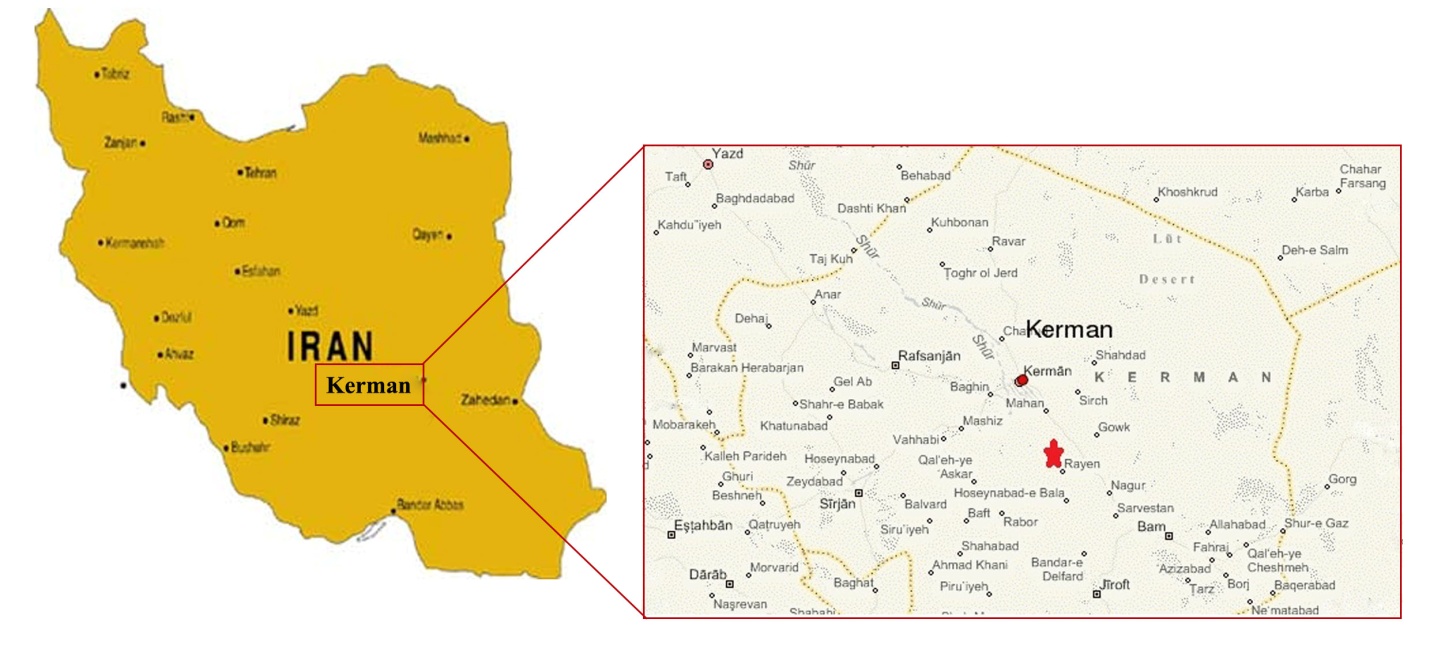 | Figure 1. Accessing roads to Bondare Bido Section, IRAN Roads ATLAS, Gitashenasi Cartographic Geographical Institute (2007) |
2.2. Lithostratigraphy of the Upper Cretaceous Sediments in Bondare Bido Section
- In thick cream-colored layer of limestone in this region with Cenomanian age is overlaid with other sedimentary series as below:- The alternation of medium - thick and cream- to yellow-colored layer of limestone with marl with thickness of 80m.- The alternation of marl with green or cream-colored limestone with thickness of 15m.- The thick layer of cream-colored limestone with thickness of 20m.- The green-colored marl with thickness of 50m.- The medium to thick and yellow- to cream-colored layers of limestone with thickness of 50m- The red sandstone with thickness of 15m.- The gray- to green-colored shales with thickness of 30m.- The red sandstone with thickness of 5m.These sediments are overlaid by Kerman conglomerate.
2.3. Evaluating of Ostracods in the Studied Section
- In this section, 13 genuses of ostracods were identified.The most famous families are Platycopida with most abundant genus of Cytherella and Cytherelloidea, Podocopida, Pontocyprella, Bairdoppilata, Pterygocythereis, Peleriops.Ostracods in Bondare Bido Section include:The ostracods in samples from lower part of this section are less in numbers. The samples of with the Upper parts of section, Campanian-Maastrichtian in age yielded good number of ostracod. The number and diversity of ostracods are low in shales and calcareous shales. The numbers of ostracods are significantly increased in marls, median shales and calcareous bands. The diversity of ostracods has been reduced by increasing depth and transgression sea water, in this region.The type of ostracod fauna indicates the depositional environment in different parts of the studied section in a way that Cytherelloidea and Haplocytheridea genuses are related to inner to middle neritic parts. Also, in most samples, the presence of Cytherelloidea genus indicates a moderate to hot climate depositional condition.Genus such as Paracypris indicates an increase in the depth and the presence of genus such as Brachycythere sp represents a reduction in the depth of the environment. The Cytherella specifies open sea with medium depth and neritic conditions of outer shelf to outer bathyal. The Paracypris and Cytherella show marine conditions with normal salinity.
 | Figure 2. The stratigraphic column of Upper Cretaceous sediments in Bondare Bido Section |
2.4. Paleoecological Studies
- The Bondare Bido Section comprises diverse assemblages of ostracods fossil. The Platycopida family is one of the most famous families with its most abundant genuses of Cytherelloidea and Cytherella which has been found. In addition, genus of Pterygocythereis, Pontocyprella and Bairdoppilata are the abundant fossils of the platycopida family. Meanwhile, genus Cytherella with 29% and genus Cytherelloidea with 24% are the most abundant genus. The paleoecological studies are by samples were counted and percentages were done and their graphs were drawn. Carried out by counting the numbers of species and graphs of their percentage is plotted (Fig.3 and 4).
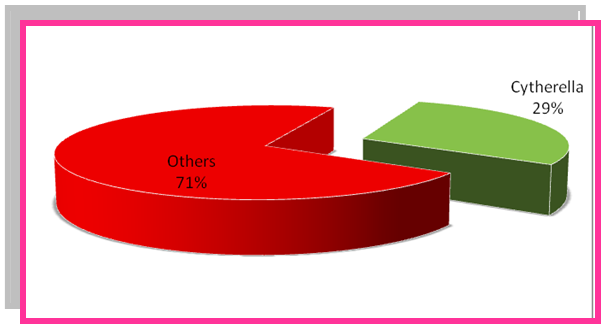 | Figure 3. The frequency percentage of Cytherella in Bondare Bido Section |
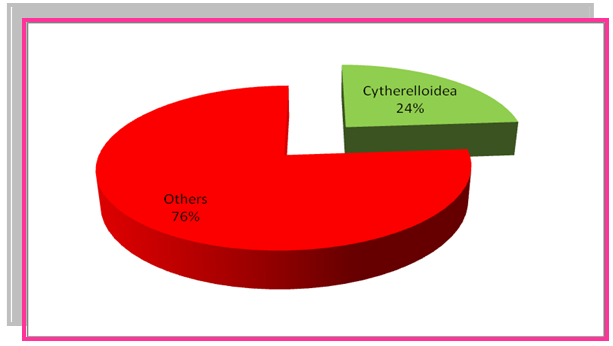 | Figure 4. The frequency percentage of Cytherelloidea in Bondare Bido Section |
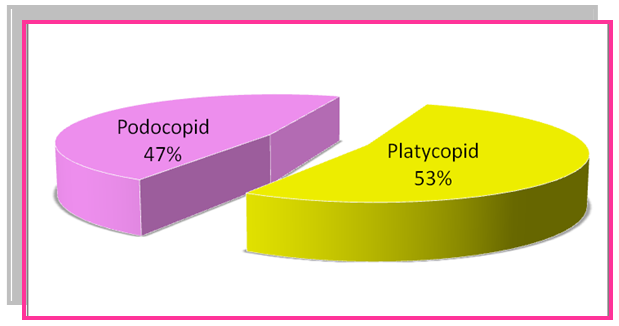 | Figure 5. Frequency percentage of Podocopid and Platycopid in Bondare Bido Section |
|
3. Temperature
- Most of the ostracods have the capability of enduring certain range of temperature. Like most of the other creatures, the ostracod assemblages are more diverse in shallow waters of low geographical latitudes than fossil ostracods belong to high latitudes (Friedwald and Mostafavi, 1998). Since the identified medium depth environment, therefore they have been benefited by proper light and temperature. The frequency of Cytherelloidea species indicate warm climatic conditions during Upper Cretaceous for Bondare Bido Section
4. Conclusions
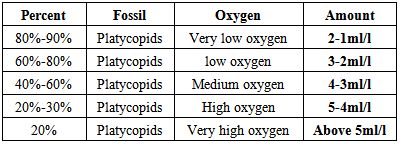
- In assessing and studying the fossil assemblages of Bondare Bido Section, 13 genus of ostracods were identified. Based on the identified planktonic foraminifera and ostracods, in Bondare Bido Section, the time interval of Upper Cretaceous sediments is proposed as from Turonian to Maastrichtian.The ostracod genus such as Haplocytheridea of Cretaceous sediments in Bondare Bido Section, may indicate marine environments with medium salinity of 36 to 39 per thousand. On the bases of identified ostracod assemblages of studied section, it can be said that during deposition of this formation, the salinity of sedimentary basin was 30 to 40 per thousand.The increase of fossil ostracod of platycopid families show the reduction of the oxygen of the environment by (Whatley et al, 2003). With regard to the presence of platycopids in the studied section which is 40 to 60%, and it can be concluded that during the deposition of Upper Cretaceous sediments in Bondare Bido Section, the environment had medium oxygen.The presence of Cytherelloidea, Bairdoppilata and Pontocyprella, indicate deep sea environment and along with Brachycythere, this ostracod fossil assemblages indicate the deep part of the platform.Since the identified medium depth environment, therefore they have been benefited by proper light and temperature. The frequency of Cytherelloidea species indicate warm climatic conditions during Upper Cretaceous for Bondare Bido Section.
Appendix
 | Plate 1. |
 | Plate 2. |
 | Plate 3. |
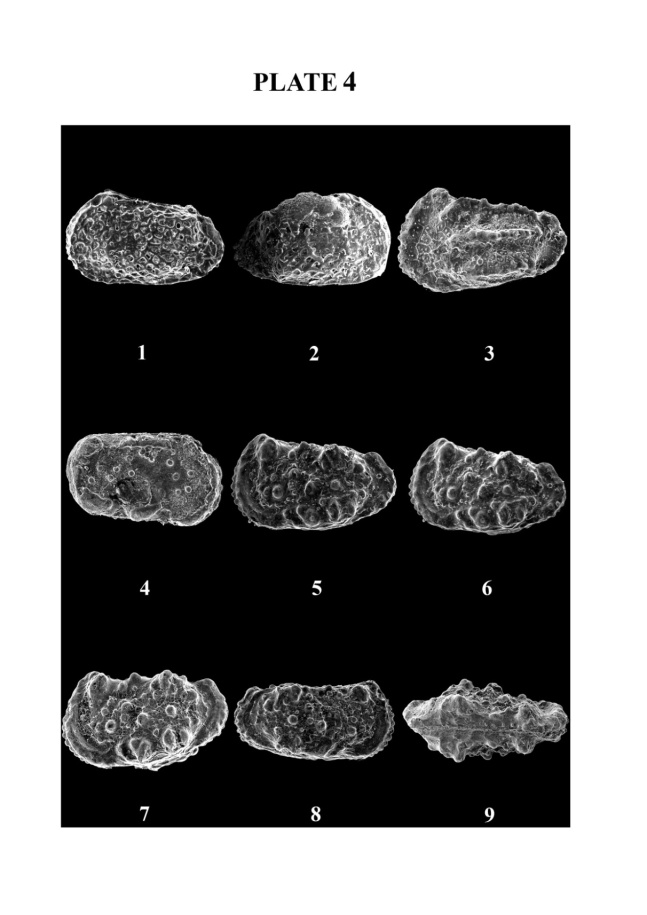 | Plate 4. |
 | Plate 5. |
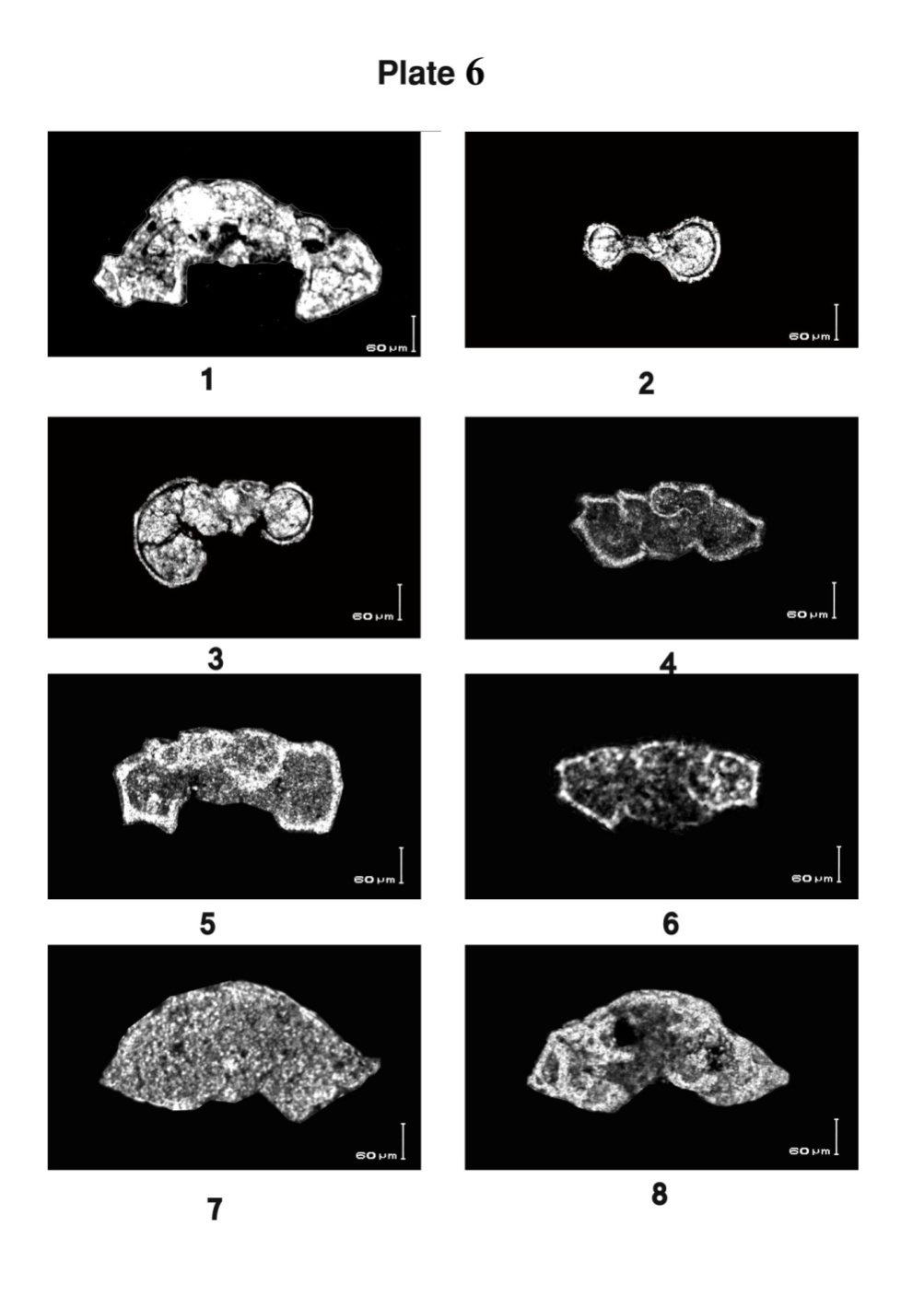 | Plate 6. |