| [1] | A. Carvalheira, et.al., (2016). Enrichment of Acinetobacter spp. from food samples. |
| [2] | A. Carvalheira, et.al., (2017). Prevalence and antimicrobial susceptibility of Acinetobacter spp. isolated from meat. Int J Food Microbiol, 243, pp. 58-63 http://dx.doi.org/10.1016/j.ijfoodmicro.2016.12.001. |
| [3] | Gonzalez-Villoria, A.M and Valverde-Garduno, V. (2016). “Antibiotic-resistant Acinetobacter baumannii increasing success remains a challenge as a nosocomial Pathogen,” Journal of Pathogens, vol. 2016, Article ID 7318075, pp. 1–10. |
| [4] | A.Y. Peleg, et.al., (2008). Acinetobacter baumannii: emergence of a successful pathogen. |
| [5] | Abbott, I., Cerqueira, G.M., Bhuiyan, S., Peleg, A.Y. (2013). Carbapenem resistance in Acinetobacter baumannii: laboratory challenges, mechanistic insights, and therapeutic strategies. Expert Rev Anti-Infect Therapeutic, 11(4): 395–409. |
| [6] | Admas, A., Gelaw, B., BelayTessema, Worku, A., & Melese, A. (2020). Proportion of bacterial isolates, their antimicrobial susceptibility profile and factors associated with puerperal sepsis among post-partum/aborted women at a referral Hospital in Bahir Dar, Northwest Ethiopia. Antimicrobial resistance and infection control, 9, 14. https://doi.org/10.1186/s13756-019-0676-2 Ethiopia. Antimicrob Resist Infect Control 9, 14 (2020). https://doi.org/10.1186/s13756-019-0676-2. |
| [7] | Al Bshabshe, A., Joseph, M.R., Al Hussein, A., Haimour, W., Hamid, M.E. (2016). Multidrug resistance Acinetobacter species at the intensive care unit, Asian Pacific journal of tropical medicine. A seer Central Hospital, Saudi Arabia, A one-year analysis, pp. 903–908. S. Zhang, C. Zhu, J. K. O. Sin, and P. K. T. Mok, “A novel ultrathin elevated channel low-temperature poly-Si TFT,” IEEE Electron Device Lett., vol. 20, pp. 569–571, Nov. 1999. |
| [8] | Al-Agamy, M.H., Shibl, A.M., Ali, M.S., Khubnani, H., Radwan, H.H., Livermore, D.M. (2014). Distribution of β-lactamases in carbapenem-non-susceptible Acinetobacter baumannii in Riyadh, Saudi Arabia. J Glob Antimicrobe Resist, 2(1), 17–21. |
| [9] | Al-Hassan, L., El Mehallawy, H., Amyes, S. (2013). Diversity in Acinetobacter baumannii isolates from pediatric cancer patients in Egypt. Clin Microbiol Infect, 19(11), 1082–8. |
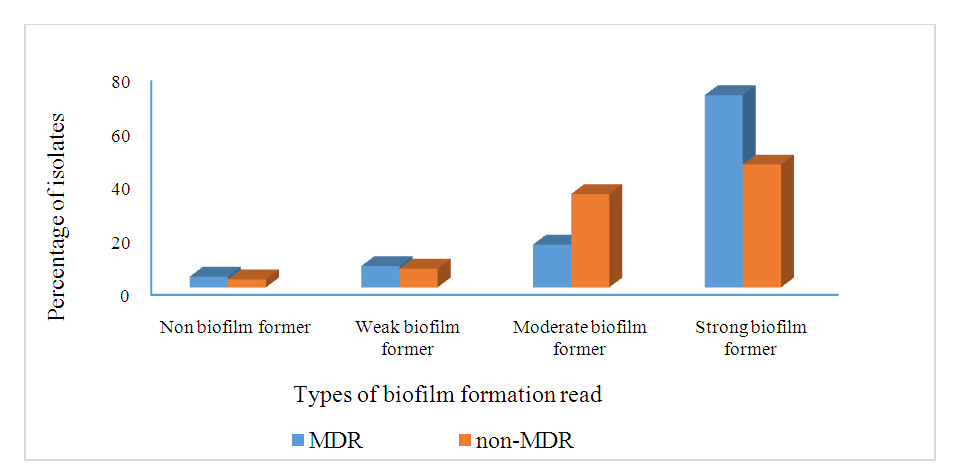
| [10] | Babapour, E., Haddadi, A., Mirnejad, R., Angaji, S.A., Amirmozafari, N. (2016). Biofilm formation in clinical isolates of nosocomial Acinetobacter baumannii and its relationship with multidrug resistance. Asian Pac J Trop Biomed, 6(6), 528–33. |
| [11] | Bradford, P. A. (2001). Extended-spectrum beta-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 14, 933–951. doi: 10.1128/cmr.14.4.933-951.2001. |
| [12] | Bush, K., et.al., (1995). A functional classification scheme for beta-lactamases and its correlation with molecular structure. Antimicrobe. Agents Chemother. 39, 1211–1233. doi: 10.1128/AAC.39.6.1211. |
| [13] | Cai, Y., Chai, D., Wang, R., Liang, B., Bai, N. (2012). Colistin resistance of Acinetobacter baumannii: clinical reports, mechanisms, and antimicrobial strategies. J. Antimicrob. Chemother, 67, 1607–1615. doi: 10.1093/jac/dks084. |
| [14] | CDC Taiwan (2016). Center for Disease Control, T. Annual report of nosocomial infections surveillance system http://www.cdc.gov.tw/english/ (2016). |
| [15] | Cicek, A.C., Saral, A., Iraz, M., Ceylan, A., Duzgun, A., Peleg, A. (2014). OXA-and GES-type β-lactamases predominate in extensively drug-resistant Acinetobacter baumannii isolates from a Turkish University hospital. Clin Microbiol Infect, 20(5), 410–5. Clin Microbiol Rev, 21 (2008), pp. 538-582. http://dx.doi.org/10.1128/CMR.00058-07. |
| [16] | Daryanavard, R., Safaei, H.R., (2015). Virulence genes and antimicrobial resistance properties of Acinetobacter baumannii isolated from pediatrics suffered from UTIs. Int. J. Adv. Res. Biol. Sci. 2(11), 272–279. |
| [17] | Durante-Mangoni, E., (2011). Global spread of drug-resistant Acinetobacter baumannii: molecular epidemiology and management of antimicrobial resistance. Future Microbiol. 6, 407–422. doi: 10.2217/fmb.11.23. |
| [18] | Evans, B. A., and Amyes, S. G. (2014). OXA beta-lactamases. Clin. Microbiol. Rev. 27, 241–263. doi: 10.1128/cmr.00117-113. |
| [19] | Fattouh, M., El-din, A.N. (2014). Emergence of carbapenem resistant Acinetobacter baumannii in the intensive care unit in Suhag University hospital, Egypt. Int J Curr Microbiol App Sci, 3(4), 732–44. |
| [20] | Moges, F., Endris, M., Mulu, A., Tessema, B., Belyhun, Y., Shiferaw, Y., Huruy, K., Unakal, C., & Kassu, A. (2014). The growing challenges of antibacterial drug resistance in Ethiopia. Journal of global antimicrobial resistance, 2(3), 148–154. https://doi.org/10.1016/j.jgar.2014.02.004. |
| [21] | Feleke, T., Eshetie, S., Dagnew, M. et al. (2018). Multidrug-resistant bacterial isolates from patients suspected of nosocomial infections at the University of Gondar Comprehensive Specialized Hospital, Northwest Ethiopia. BMC Res Notes 11, 602. Food Microbiol, 55 (2016), pp. 123-127. http://dx.doi.org/10.1016/j.fm.2015.11.002. |
| [22] | Fouad, M., Attia, A.S., Tawakkol, W.M., Hashem, A.M. (2013). Emergence of carbapenem-resistant Acinetobacter baumannii harboring the OXA-23 carbapenemase in intensive care units of Egyptian hospitals. Int J Infect Dis, 17(12), e1252–e4. |
| [23] | Ghasemi, E., Ghalavand, Z., Goudarzi, H., Yeganeh. F., Hashemi, A., Dabiri, H., Mirsamadi, E.S., Foroumand, M. (2018). Phenotypic and Genotypic Investigation of Biofilm Formation in Clinical and Environmental Isolates of Acinetobacter baumannii. Arch Clin Infect Dis, 13(4), e12914. |
| [24] | Guchi, B., Ashenafi, M. (2010). Microbial load, prevalence and antibiograms of salmonella and Shigella in lettuce and green peppers. Ethiopian Journal of Health Sciences. Mar; 20(1, 41-48. DOI: 10.4314/ejhs. v20i1.69431. |
| [25] | Chang, Y., Luan, G., Xu, Y., Wang, Y., Shen, M., Zhang, C., Zheng, W., Huang, J., Yang, J., Jia, X., & Ling, B. (2015). Characterization of carbapenem-resistant Acinetobacter baumannii isolates in a Chinese teaching hospital. Frontiers in microbiology, 6, 910. https://doi.org/10.3389/fmicb.2015.00910. |
| [26] | Lee, K., Yong, D., Jeong, S.H., Chong, Y. (2011). Multidrug-resistant Acinetobacter spp.: increasingly problematic nosocomial pathogens. Yonsei Med J. 52(6), 879–91. |
| [27] | Eveillard, M., Kempf, M., Belmonte, O., Pailhoriès, H., & Joly-Guillou, M. L. (2013). Reservoirs of Acinetobacter baumannii outside the hospital and potential involvement in emerging human community-acquired infections. International journal of infectious diseases: IJID: official publication of the International Society for Infectious Diseases, 17(10), e802–e805. https://doi.org/10.1016/j.ijid.2013.03.021. |
| [28] | Kempf, M., Rolain, J. M., Diatta, G., Azza, S., Samb, B., Mediannikov, O., Gassama Sow, A., Diene, S. M., Fenollar, F., & Raoult, D. (2012). Carbapenem resistance and Acinetobacter baumannii in Senegal: the paradigm of a common phenomenon in natural reservoirs. PloS one, 7(6), e39495. https://doi.org/10.1371/journal.pone.0039495. |
| [29] | Kempf, M., & Rolain, J. M. (2012). Emergence of resistance to carbapenems in Acinetobacter baumannii in Europe: clinical impact and therapeutic options. International journal of antimicrobial agents, 39(2), 105–114. https://doi.org/10.1016/j.ijantimicag.2011.10.004. |
| [30] | Lin, M. F., & Lan, C. Y. (2014). Antimicrobial resistance in Acinetobacter baumannii: From bench to bedside. World journal of clinical cases, 2(12), 787–814. https://doi.org/10.12998/wjcc.v2.i12.787. |
| [31] | Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., Harbarth, S., Hindler, J. F., Kahlmeter, G., Olsson-Liljequist, B., Paterson, D. L., Rice, L. B., Stelling, J., Struelens, M. J., Vatopoulos, A., Weber, J. T., & Monnet, D. L. (2012). Multidrug-resistant, extensively drug-resistant, and pan drug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clinical microbiology and infection: the official publication of the European Society of Clinical Microbiology and Infectious Diseases, 18(3), 268–281. https://doi.org/10.1111/j.1469-0691.2011.03570.x. |
| [32] | Maraki, S., Mantadakis, E., Mavromanolaki, V. E., Kofteridis, D. P., & Samonis, G. (2016). A 5-year Surveillance Study on Antimicrobial Resistance of Acinetobacter baumannii Clinical Isolates from a Tertiary Greek Hospital. Infection & chemotherapy, 48(3), 190–198. https://doi.org/10.3947/ic.2016.48.3.190. |
| [33] | Mathlouthi, N., Ben Lamine, Y., Somai, R., Bouhalila-Besbes, S., Bakour, S., Rolain, J. M., & Chouchani, C. (2018). Incidence of OXA-23 and OXA-58 Carbapenemases Coexpressed in Clinical Isolates of Acinetobacter baumannii in Tunisia. Microbial drug resistance (Larchmont, N.Y.), 24(2), 136–141. https://doi.org/10.1089/mdr.2016.0306. |
| [34] | Belyhun, Y., Moges, F., Endris, M. (2018). Ocular bacterial infections and antibiotic resistance patterns in patients attending Gondar Teaching Hospital, Northwest Ethiopia. BMC Res Notes 11, 597. https://doi.org/10.1186/s13104-018-3705-y. |
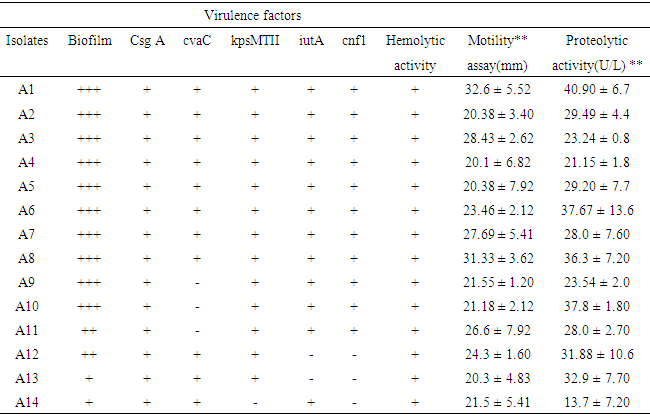
| [35] | Mohajeri, P., Sharbati, S., Farahani, A., & Rezaei, Z. (2016). Evaluate the frequency distribution of nonadhesive virulence factors in carbapenemase-producing Acinetobacter baumannii isolated from clinical samples in Kermanshah. Journal of natural science, biology, and medicine, 7(1), 58–61. https://doi.org/10.4103/0976-9668.175071. |
| [36] | Momtaz, H., Seifati, S.M., Tavakol, M. (2015). Determining the prevalence and detection of the most prevalent virulence genes in Acinetobacter baumannii Isolated from hospital Infections. International Journal of Medical Laboratory, 2(2), 87-97. |
| [37] | Belmonte, O., Pailhoriès, H., Kempf, M., Gaultier, M. P., Lemarié, C., Ramont, C., Joly-Guillou, M. L., & Eveillard, M. (2014). High prevalence of closely related Acinetobacter baumannii in pets according to a multi-center study in veterinary clinics, Reunion Island. Veterinary-microbiology, 170(3-4), 446–450. https://doi.org/10.1016/j.vetmic.2014.01.042. |
| [38] | OMEGA Innovations in nucleic acid isolation http://2015.igem.org/wiki/images/1/18/NUDT_CHIAN-Protoco1.pdf. |
| [39] | Peleg, A., et. al., (2008). Acinetobacter baumannii: emergence of a successful pathogen. Clin. Microbiol. Rev. 21, 538–582. doi: 10.1128/cmr.00058-57. |
| [40] | Qi, L., Li, H., Zhang, C., Liang, B., Li, J., Wang, L., Du, X., Liu, X., Qiu, S., & Song, H. (2016). Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Frontiers in microbiology, 7, 483. https://doi.org/10.3389/fmicb.2016.00483. |
| [41] | Rafei, R., et al. (2015). Extra-human epidemiology of Acinetobacter baumannii in Lebanon. Appl Environ Microbiol, 81, pp. 2359-2367. http://dx.doi.org/10.1128/AEM.03824-14. |
| [42] | Richard D. et.al., (1994). A Phylogenetic Tree of 16S rRNA Sequences from Sulfate Reducing Bacteria in a Sandy Marine Sediment. Applied and environmental Microbiology, Sept. 1994, p. 3437-3439, Vol. 60, No. |
| [43] | Richet, H. M., Mohammed, J., McDonald, L. C., & Jarvis, W. R. (2001). Building communication networks: international network for the study and prevention of emerging antimicrobial resistance. Emerging infectious diseases, 7(2), 319–322. https://doi.org/10.3201/eid0702.010235. |
| [44] | Thummeepak, R., Kongthai, P., Leungtongkam, U., & Sitthisak, S. (2016). Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. International microbiology: the official journal of the Spanish Society for Microbiology, 19(2), 121–129. https://doi.org/10.2436/20.1501.01.270. |
| [45] | Tiwari V., et. al. (2012). Carbapenem-hydrolyzing oxacillinase in high resistant strains of Acinetobacter baumannii isolated from India. Microb. Pathog. 53, 81–86. doi: 10.1016/j.micpath.2012.05.004. |
| [46] | Walther-Rasmussen, J., et.al. (2006). OXA-type carbapenemases. J. Antimicrobe. Chemother. 57, 373–383. doi: 10.1093/jac/dki482. |
| [47] | Yaw Anane A., et.al., (2019). Prevalence and molecular analysis of multidrug-resistant Acinetobacter baumannii in the extra-hospital environment in Mthatha, South Africa. The Brazilian journal of infectious diseases. Vol. 23. Issue 6. pages 371-380 (Nov. – Dec. 2019). |
| [48] | Youn Sung J. (2018). Molecular Characterization and Antimicrobial Susceptibility of Biofilm-forming Acinetobacter baumannii Clinical Isolates from Daejeon, Korea, vol. 50. |
| [49] | Zeighami, H., Valadkhani, F., Shapouri, R. (2019). Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect Dis 19, 629. https://doi.org/10.1186/s12879-019-4272-0. |
| [50] | Asmaru, G. , Samuel S. (2013). Microbial Spectrum of Fruit in Gondar town Markets, Northwestern Ethiopia, Journal of Microbiology Research, 3(1): 1-10. doi:10.5923/j.microbiology.20130301.01. |
| [51] | Akoachere, F. T., Bertrand, F., and Joseph, M. N. (2018). Bacterial and parasitic contaminants of salad vegetables sold in markets in Fako Division, Cameroon and evaluation of hygiene and handling practices of vendors. BMC Research Notes, 11, 100. https://doi.org/10.1186/s13104-018-3175-2. |
| [52] | Bennett, R. L., French, K. S., Resta, R. G., & Doyle, D. L. (2008). Standardized human pedigree nomenclature: update and assessment of the recommendations of the National Society of Genetic Counselors. Journal of genetic counseling, 17(5), 424–433. https://doi.org/10.1007/s10897-008-9169-9. |
| [53] | Ragupathi, N., Bakthavatchalam, Y. D., Mathur, P., Pragasam, A. K., Walia, K., Ohri, V. C., & Veeraraghavan, B. (2019). Plasmid profiles among some ESKAPE pathogens in a tertiary care centre in south India. The Indian journal of medical research, 149(2), 222–231. https://doi.org/10.4103/ijmr.IJMR_2098_17. |
| [54] | Santajit, S., & Indrawattana, N. (2016). Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed research international, 2016, 2475067. https://doi.org/10.1155/2016/2475067. |
| [55] | Bergogne-Bérézin, E., & Towner, K. J. (1996). Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clinical microbiology reviews, 9(2), 148–165. https://doi.org/10.1128/CMR.9.2.148. |
| [56] | McConnell, M. J., Actis, L., & Pachón, J. (2013). Acinetobacter baumannii: human infections, factors contributing to pathogenesis and animal models. FEMS microbiology reviews, 37(2), 130–155. https://doi.org/10.1111/j.1574-6976.2012.00344. |
| [57] | Hrenovic, J., Durn, G., Goic-Barisic, I., & Kovacic, A. (2014). Occurrence of an environmental Acinetobacter baumannii strain similar to a clinical isolate in paleosol from Croatia. Applied and environmental microbiology, 80(9), 2860–2866. https://doi.org/10.1128/AEM.00312-14. |
| [58] | Hrenovic, J., Ganjto, M., Goic-Barisic. I. (2017). "Carbapenem-resistant bacteria in a secondary wastewater treatment plant. Gale Academic OneFile, " Water SA, vol. 43, no. 2, Apr. 2017, pp. 186. |
| [59] | Dijkshoorn, L., Nemec, A. & Seifert, H. (2007). An increasing threat in hospitals: multidrug-resistant Acinetobacter-baumannii. Nat-Rev-Microbiol 5, 939–951 https://doi.org/10.1038/nrmicro1789. |
| [60] | Roca, I., Espinal, P., Vila-Farrés, X., & Vila, J. (2012). The Acinetobacter baumannii Oxymoron: Commensal Hospital Dweller Turned Pan-Drug-Resistant Menace. Frontiers in microbiology, 3, 148. https://doi.org/10.3389/fmicb.2012.00148. |
| [61] | Vázquez-López, R., Solano-Gálvez, S. G., Juárez Vignon-Whaley, J. J., Abello Vaamonde, J. A., Padró Alonzo, L. A., Rivera Reséndiz, A., Muleiro Álvarez, M., Vega López, E. N., Franyuti-Kelly, G., Álvarez-Hernández, D. A., Moncaleano Guzmán, V., Juárez Bañuelos, J. E., Marcos Felix, J., González Barrios, J. A., & Barrientos Fortes, T. (2020). Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics (Basel, Switzerland), 9(4), 205. https://doi.org/10.3390/antibiotics9040205. |
| [62] | Manchanda, V., Sanchaita, S., & Singh, N. (2010). Multidrug resistant acinetobacter. Journal of global infectious diseases, 2(3), 291–304. https://doi.org/10.4103/0974-777X.68538. |
| [63] | Tooke, C. L., Hinchliffe, P., Bragginton, E. C., Colenso, C. K., Hirvonen, V., Takebayashi, Y., & Spencer, J. (2019). β-Lactamases and β-Lactamase Inhibitors in the 21st Century. Journal of molecular biology, 431(18), 3472–3500. https://doi.org/10.1016/j.jmb.2019.04.002. |
| [64] | Drawz, S. M., & Bonomo, R. A. (2010). Three decades of beta-lactamase inhibitors. Clinical microbiology reviews, 23(1), 160–201. https://doi.org/10.1128/CMR.00037-09. |
| [65] | Page, M. I., & Badarau, A. (2008). The mechanisms of catalysis by metallo beta-lactamases. Bioinorganic-chemistry-and-applications, 2008, 576297. https://doi.org/10.1155/2008/576297. |
| [66] | Papp-Wallace, K. M., Endimiani, A., Taracila, M. A., & Bonomo, R. A. (2011). Carbapenems: past, present, and future. Antimicrobial agents and chemotherapy, 55(11), 4943–4960. https://doi.org/10.1128/AAC.00296-11. |
| [67] | Hammoudi Halat, D., & Ayoub Moubareck, C. (2020). The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics (Basel, Switzerland), 9(4), 186. https://doi.org/10.3390/antibiotics9040186. |
| [68] | Senkyrikova, M., Husickova, V., Chroma, M., Sauer, P., Bardon, J., & Kolar, M. (2013). Acinetobacter baumannii producing OXA-23 detected in the Czech Republic. Springer Plus, 2(1), 296. https://doi.org/10.1186/2193-1801-2-296. |
| [69] | Asif, M., Alvi, I. A., & Rehman, S. U. (2018). Insight into Acinetobacter baumannii: pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infection and drug resistance, 11, 1249–1260. https://doi.org/10.2147/IDR.S166750. |
| [70] | Manchanda, V., Sanchaita, S., & Singh, N. (2010). Multidrug resistant acinetobacter. Journal of global infectious diseases, 2(3), 291–304. https://doi.org/10.4103/0974-777X.68538. |
| [71] | Askari, N., Momtaz, H., & Tajbakhsh, E. (2020). Prevalence and phenotypic pattern of antibiotic resistance of Acinetobacter baumannii isolated from different types of raw meat samples in Isfahan, Iran. Veterinary medicine and science, 6(1), 147–153. https://doi.org/10.1002/vms3.199. |
| [72] | World Health Organization, 2007. A safer future. Global Public Health Security in the 21st Century. https://www.who.int/whr/2007/whr07_en.pdf. |
| [73] | Akinde1, S. B., Abiodun A. Sunday, A. A., Folasade M. Adeyemi,F. M., Iyabobola B. Fakayode1, I. B, Odunola O. Oluwajide, O.O., Adetoun A. Adebunmi1, A. A., Julius K. Oloke, J. K., and Clement O. Adebooye. C.O. (2016). Microbes in Irrigation Water and Fresh Vegetables: Potential Pathogenic Bacteria Assessment and Implications for Food Safety, Applied Biosafety: Journal of ABSA International 2016, Vol. 21(2) 89-97. |
| [74] | Venkatesan, M. M., Goldberg, M. B., Rose, D. J., Grotbeck, E. J., Burland, V., & Blattner, F. R. (2001). Complete DNA sequence and analysis of the large virulence plasmid of Shigella flexneri. Infection and immunity, 69(5), 3271–3285. https://doi.org/10.1128/IAI.69.5.3271-3285.2001. |
| [75] | Clarridge J. E., 3rd (2004). Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clinical microbiology reviews, 17(4), 840–862. https://doi.org/10.1128/CMR.17.4.840-862.2004. |
| [76] | Janda, J. M., & Abbott, S. L. (2007). 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: pluses, perils, and pitfalls. Journal of clinical microbiology, 45(9), 2761–2764. https://doi.org/10.1128/JCM.01228-07. |
| [77] | Zalts, R., Neuberger, A., Hussein, K., Raz-Pasteur, A., Geffen, Y., Mashiach, T., & Finkelstein, R. (2016). Treatment of Carbapenem-Resistant Acinetobacter baumannii Ventilator-Associated Pneumonia: Retrospective Comparison Between Intravenous Colistin and Intravenous Ampicillin-Sulbactam. American journal of therapeutics, 23(1), e78–e85. https://doi.org/10.1097/MJT.0b013e3182a32df3. |
| [78] | Bahador, A., Raoofian, R., Pourakbari, B., Taheri, M., Hashemizadeh, Z., & Hashemi, F. B. (2015). Genotypic and Antimicrobial Susceptibility of Carbapenem-resistant Acinetobacter baumannii: Analysis of is Aba Elements and bla OXA-23-like Genes Including a New Variant. Frontiers in microbiology, 6, 1249. https://doi.org/10.3389/fmicb.2015.01249. |
| [79] | Al-Kadmy, I., Ali, A., Salman, I., & Khazaal, S. S. (2017). Molecular characterization of Acinetobacter baumannii isolated from Iraqi hospital environment. New microbes and new infections, 21, 51–57. https://doi.org/10.1016/j.nmni.2017.10.010. |
| [80] | Hassan, A., Usman, J., Kaleem, F., Omair, M., Khalid, A., & Iqbal, M. (2011). Evaluation of different detection methods of biofilm formation in the clinical isolates. The Brazilian journal of infectious diseases: an official publication of the Brazilian Society of Infectious Diseases, 15(4), 305–311. |
| [81] | Abdel Halim, R. M., Kassem, N. N., & Mahmoud, B. S. (2018). Detection of Biofilm Producing Staphylococci among Different Clinical Isolates and Its Relation to Methicillin Susceptibility. Open access Macedonian journal of medical sciences, 6(8), 1335–1341. https://doi.org/10.3889/oamjms.2018.246. |
| [82] | Antunes, L. C., Imperi, F., Carattoli, A., & Visca, P. (2011). Deciphering the multifactorial nature of Acinetobacter baumannii pathogenicity. PloS one, 6(8), e22674. https://doi.org/10.1371/journal.pone.0022674. |
| [83] | Avishai Ben-Davida, Charles E. Davidson, (2014). Estimation method for serial dilution experiments, Journal of Microbiological Methods, Volume 107, December 2014, Pages 214-22. https://doi.org/10.1016/j.mimet.2014.08.023. |
| [84] | Tsegaye Alemayehu, (2020). Prevalence of multidrug-resistant bacteria in Ethiopia: a systematic review and meta-analysis, Journal of Global Antimicrobial Resistance Volume 26, September 2021, Pages 133-139. https://doi.org/10.1016/j.jgar.2021.05.017. |
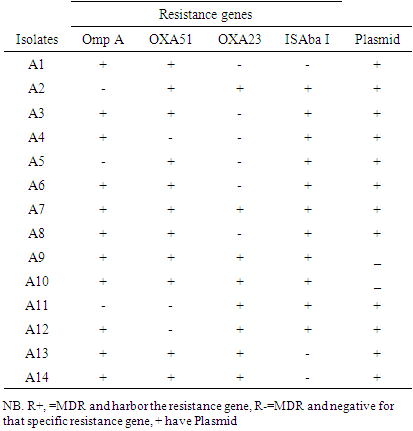
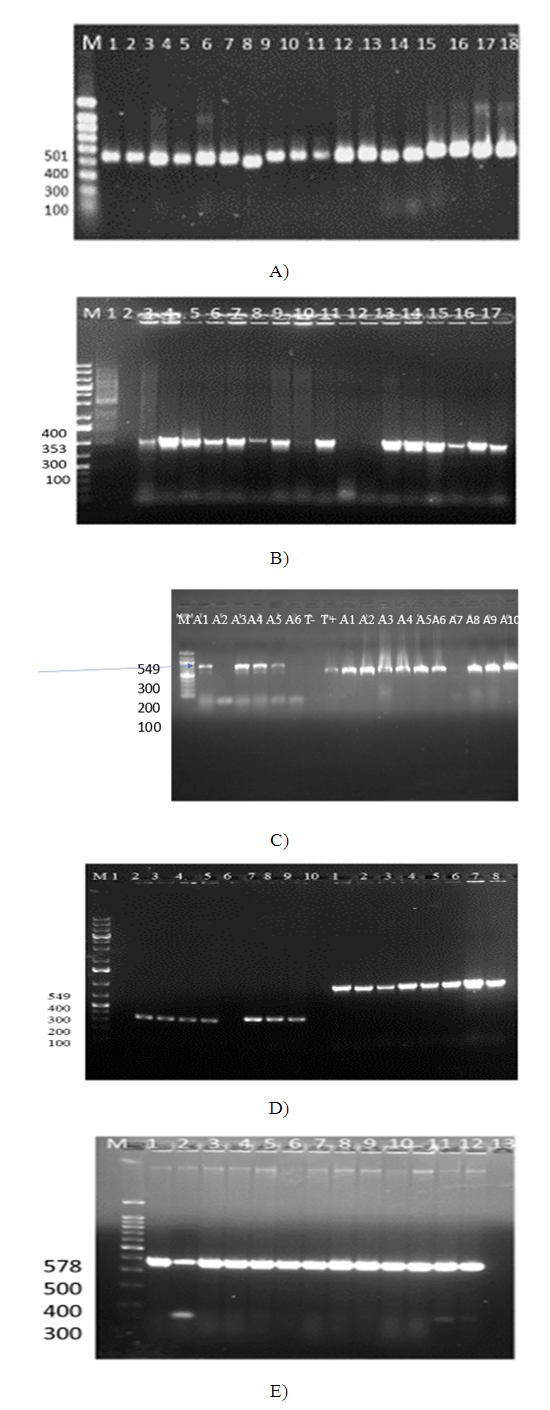
| [85] | Turton, J. F., Woodford, N., Glover, J., Yarde, S., Kaufmann, M. E., & Pitt, T. L. (2006). Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. Journal of clinical microbiology, 44(8), 2974–2976. https://doi.org/10.1128/JCM.01021-06. |
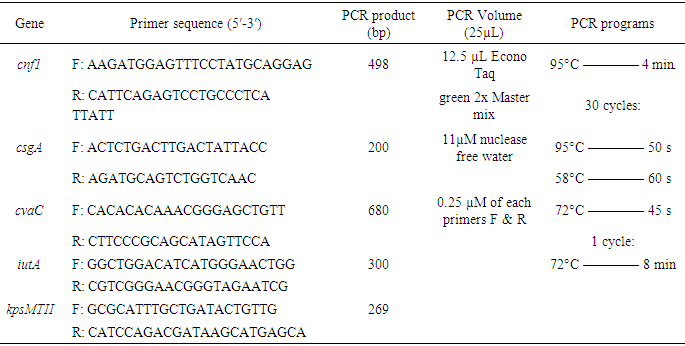
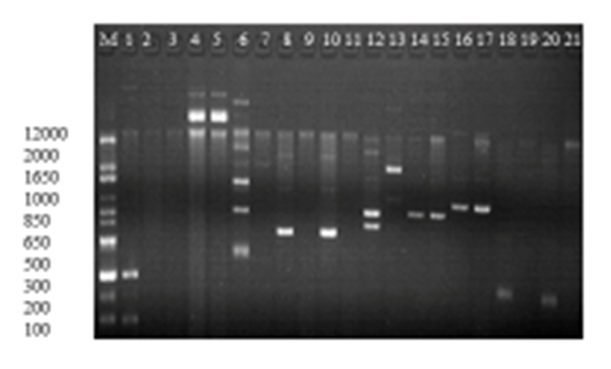
| [86] | Tavakol, M., Momtaz, H., Mohajeri, P. et al. Genotyping and distribution of putative virulence factors and antibiotic resistance genes of Acinetobacter baumannii strains isolated from raw meat. Antimicrob Resist Infect Control 7, 120 (2018). https://doi.org/10.1186/s13756-018-0405-2R. E. Sorace, V. S. Reinhardt, and S. A. Vaughn, “High-speed digital-to-RF converter,” U.S. Patent 5 668 842, Sept. 16, 1997. |

 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML