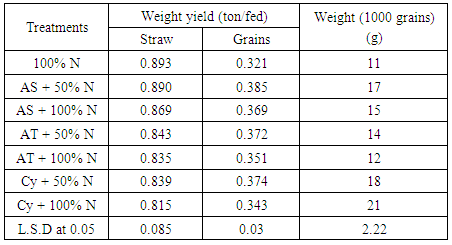
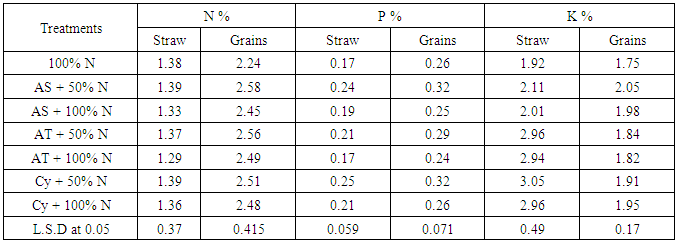
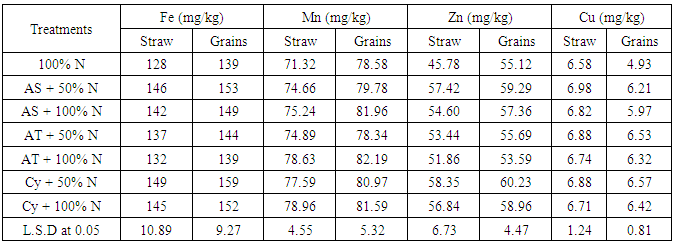
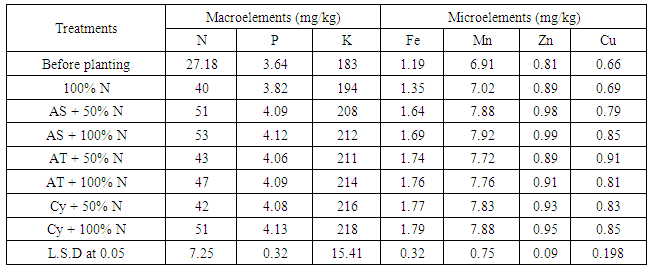
| [1] | Shen X., Hu H., Peng H., Wang W. and Zhang X. (2013). Comparative genomic analysis of four representative plant growth promoting rhizobacteria in Pseudomonas. BMC Genomics.14: 271. |
| [2] | Deshwal V.K. and Kumar P. (2013). Production of Plant growth promoting substance by Pseudomonads. J. Acad. Indus. Res. 2(4): 221-225. |
| [3] | UNEP (2009). The environmental food crisis: The environments’ role in averting future food crisis. |
| [4] | Gehad A. (2003). Deteriorated Soils in Egypt: Management and Rehabilitation. (Extent and causes of salt affected soils). Executive authority for land improvement projects (EALIP), Arab Republic of Egypt, Ministry of Agriculture and Land Reclamation, pp 15-19. |
| [5] | Han H.S. and Lee K.D. (2005). Plant Growth Promoting Rhizobacteria effect on antioxidant status, photosynthesis, mineral uptake and growth of lettuce under soil salinity. Res. J. Agri. Biol.Sci. 1(3): 210-215. |
| [6] | Shukla P.S., Agarwal P.K. and Jha B. (2012). Improved salinity tolerance of Arachis hypogaea (L.) by the Interaction of halotolerant plant-growth-promoting rhizobacteria. J. Plant Growth Regul. 31(2): 195-206. |
| [7] | Upadhyay S.K., Singh J.S. and Singh D.P. (2011). Exopolysaccharide-Producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere. 21(2): 214-222. |
| [8] | Egamberdieva D. (2009). Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol Plant, 31:861–864. |
| [9] | Chelikani P., Fita I. and Loewen P.C. (2004). Diversity of structures and properties among catalases. Cell. Mol. Life Sci. 61(2): 192–208. |
| [10] | Kalir A.M. and Poljakoff-Mayber A. (1981). Changes in Activity of Malate Dehydrogenase, Catalase, Peroxidase and Superoxide Dismutase in Leaves of Halimione portulacoides (L.) Aellen Exposed to High Sodium Chloride Concentrations. Ann. Bot. 47(1): 75-85. |
| [11] | Saraf N. (2013). Enhancement of Catalase Activity under Salt Stress in Germinating Seeds of Vigna radiata. Asian Journal of Biomedical and Pharmaceutical Sciences 3(17): 6-8. |
| [12] | Howitt M.S. and Udvardi M.K. (2000). Structure, function and regulation of ammonium transporters in plants. Biochim. Biophys 152-170. |
| [13] | Dobereiner J., Marriel I.E. and Nery M. (1976). Ecological distribution of Spirillum lipoferum Beijerinck. Canad. J. Microbiol. 22: 1464-1473. |
| [14] | Hegazy N.A. and Neimela S. (1976). Anote on the astimation of Azotobacter densities by membrane filter technique. J. Appl. Bacteriol. 41: 311. |
| [15] | Venkataraman G.S. (1972). Algal Biofertilizer and Rice Cultivation. Today and Tomorrow’s Printer and Publishers, New Delhi. |
| [16] | Holt J.G., Krieg N.R., Sneath P.H.A., Staley J.T. and Williamsn S.T. (1994). Group 11. Oxygenic phototrophic bacteria. In: Bergey's Manual of Determinative Bacteriology, 9th edn (ed. Holt JG), pp. 377–425. Williams and Wilkins, Baltimore. |
| [17] | Sergeeva E., Liaimer A. and Bergman B. (2002). Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta 215: 229-238. |
| [18] | Sabra W., Zeng A.P., Lünsdorf H. and Deckwer W.D. (2000). Effect of oxygen on formation and structure of Azotobacter vinelandii alginate and its role in protecting nitrogenase. Appl. Environ. Microbiol. 66: 4037–4044. |
| [19] | Kaushik A.A., Nisha R. and Kaushik C.P. (2010). Effect of cyanobacteria lexopolysaccharides on salt stress alleviation and seed germination. J. Environm.l Biol., 31(5): 701-704. |
| [20] | Cappuccino J.C. and Sherman N. (1992). In: Microbiology: A Laboratory Manual, third ed. Benjamin/cummings Pub. Co. New York, pp. 125–179. |
| [21] | Jackson M.L. (1973). Soil Chemical Analysis. pr. entice Hall Inc., N.J. 498pp. |
| [22] | Casida L.E., Klein D.A. and Suntoro T. (1964). Soil dehydrogenase activity. Soil Sci. 98: 371-376. |
| [23] | LethBridage G., Davidson M.S. and Sparling G.P. (1982). Critical evaluation of the acetylene reduction test for estimating the nitrogenase activity of nitrogen fixing bacteria associated with the roots of wheat and barley. Soil Biol. and Biochem. 14: 27-35. |
| [24] | Page A.I., Miller R.H. and Keeney D.R. (1982). Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties. 2nd Edition, Amer. Soc. of Agron., Madison, Wisconsin, U.S.A. |
| [25] | Cottenie A., Verloo M., Kiekens L., Velghe G. and Camerlynck R. (1982). Chemical Analysis of Plants and Soils. Laboratory of Analytical and Agrochemistry, State University-Ghent, Belgium. |
| [26] | Sendecore G.W. and Cochran N.G. (1980). Statistical methods. 7th edition. Iowa State University. Press. Ames. |
| [27] | Noumavo P.A., Agbodjato N.A., Gachomo E.W., Salami H.A., Baba-Moussa F., Adjanohoun A., Kotchoni S.G. and Baba-Moussa L. (2015). Metabolic and biofungicidal properties of maize rhizobacteria for growth promotion and plant disease resistance. African J. Biotech. 14(9): 811-819. |
| [28] | Srivastava A.K., Bhargava P., Kumar A., Rai L.C. and Neilan B.A. (2009). Molecular characterization and the effect of salinity on cyanobacterial diversity in the rice fields of Eastern Uttar Pradesh, India. Saline Sys. 5: 1-17. |
| [29] | Arora M., Kaushik A., Rani N. and Kaushik C.P. (2010). Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination. Journal of Environmental Biology 31(5): 701-704. |
| [30] | Ahmed A., Hasnain A., Akhtar S., Hussain A., Yasin A.U.G. and Wahid A. et al. (2010). Antioxidant enzymes as bio-markers for copper tolerance in safflower (Carthamus tinctorius L.). African Journal of Biotechnology 9(33): 5441-5444. |
| [31] | Tel-Or E., Huflejt M.E. and Packer L. (1986). Hydroperoxide metabolism in cyanobacteria. Arch Biochem Biophys. 246: 396-402. |
| [32] | Bienert, G.P.; J. K. Schjoerring, T. P. Jahn (2006): Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta (BBA) –Biomembranes, 1758 (8), 997-1003. |
| [33] | Nain L., Rana A., Joshi M., Jadhav S.D., Kumar D., Shivay Y.S., Paul S. and Prasanna R. (2010). Evaluation of synergistic effects of bacterial and cyanobacterial strains as biofertilizers for wheat. Plant and Soil, 331, (1), 217-230. |
| [34] | Abou-El-Eyoun A.T. (2005). Studies on the role of cyanobacteria in agriculture. M.Sc. Thesis, Soil Dpt. Faculty of Agriculture, Menia Univ. |
| [35] | Abou-Zeid M.Y. and Bakry M.A.A. (2011). Integrated Effect of Bio-organic Manures and Mineral Ertilizers on Potato Productivity and the Fertility Status of a Calcareous Soil. Aust. J. of Basic and Appl.Sci. 5(8): 1385-1399. |
| [36] | Swarnalakshmi K., Dhar D. and Singh P. (2007). Evaluation of blue-green algal inoculation on specific soil parameters. Acta Agronomica Hungarica 55: 307-313. |
| [37] | Prasanna R., Joshi M., Rana A., Singh Y., Nain L. (2011). Influence of co-inoculation of bacteria-cyanobacteria on crop yield and C–N sequestration in soil under rice crop. World J. Microbiol. Biotechnol. 926-929. |
| [38] | Ghazal F.M., Hassan M.M.M., EL-Sayed G.A.M. and Desoky A.H. (2013a). Response of Maize Crop to Cyanobacteria Applied Under Different Nitrogen Rates. Nature and Science 11(12): 172-181. |
| [39] | Ghazal F.M., El-Koomy M.B.A., Abdel-Kawi Kh.A. and Soliman M.M. (2013b). Impact of Cyanobacteria, Humic Acid and Nitrogen Levels on Maize (Zea mays L.) Yield and Biological Activity of the Rhizosphere in Sandy Soils. Journal of American Science 9(2): 46- 55. |
| [40] | Zayed B.A., Elkhoby W.M., Salem A.K., Ceesay M., and Uphoff N.T. (2013). Effect of Integrated Nitrogen Fertilizer on Rice Productivity and Soil Fertility under Saline Soil Conditions. J. Plant Biology Res. 2:14. |
| [41] | Saad A.A. and Mostafa Y.S. (2009). Effect of nitrogen supply and Azospirillum brasilense Sp-248 on the response of wheat to seawater irrigation. Saudi J. Biol.Scie. 16: 101–107. |
| [42] | Sartaj A.W., Chand S. and Ali T. (2013). Potential Use of Azotobacter Chroococcum in Crop Production: An Overview. Curr. Agri. Res. J. 1: 35-38. |
| [43] | Chaudhary D., Narula N., Sindhu S.S. and Behl R.K. (2013). Plant growth stimulation of wheat (Triticum aestivum L.) by inoculation of salinity tolerant Azotobacter strains. Physiol. Mol. Biol. Plants 19(4): 515-519. |
| [44] | De Philippis R. and Vincenzini M. (1998). Extracellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 22: 151-175. |
| [45] | Rifat H., Safdar A., Ummay A., Rabia Kh. and Iftikhar A. (2010). Soil beneficial bacteria and their role in plant growth promotion. J. Ann. Microbiol. 7: 117-137. |
| [46] | Perotti E.B.R. and Pidello A. (2012). Plant-Soil-Microorganism Interactions on Nitrogen Cycle: Azospirillum Inoculation. Published in Environmental Sciences. "Advances in Selected Plant Physiology Aspects", book edited by Giuseppe Montanaro and BartolomeoDichio, pg 189-208. |
| [47] | Shaban K.A., Abd El-Kader M.G. and Khalil Z.M. (2012). Effect of soil amendments on soil fertility and sesame crop productivity under newly reclaimed soil conditions Journal of Applied Sciences Research 8(3): 1568-1575. |
| [48] | Prasanna R., Bidyarani N., Babu S., Hossain F., Shivay Y.S. and Nain L. (2015). Cyanobacterial inoculation elicits plant defense response and enhanced Zn mobilization in maize hybrids. Cogent Food and Agriculture 1: 998507. |
| [49] | Tantawy E.A. and. Shaban K.A. (2010). Exopolysaccharide and indole acetic acid producing endophytic bacteria have an essential role in Rice (Oryza sativa) resistance in salt affected soils. New Egypt J. Microbial. 27: 210-227. |
| [50] | Brady N.C. and Weil R.R. (2005). The Nature and Properties of Soil (13th Ed.). Macmillan Publi. Co., New York. |

 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML