| [1] | Mayes, JS. and Watson, GH. (2004): Direct effects of sex steroid hormones on adipose tissues and obesity. Obes. Rev., 5 (4): 197-216. |
| [2] | Fernandez, c., Bellentani, F., Fernandes, G., Perobelli, J., Favareto, AP., Nascimento, A., Cicogna, A. and Kempinas, W. (2011): Diet-induced obesity in rats leads to a decrease in sperm motility. Reproductive Biology and Endocrinology, 9 (32): 1-10. |
| [3] | Jensen,TK., Andersson, AM., Jørgensen, N., Andersen, AG, Carlsen E, Petersen JH. and Skakkebæk NE. (2004): Body mass index in relation to semen quality and reproductive hormones among 1,558 Danish men. Fertil. Steril., 82 (4): 863-870. |
| [4] | Hammoud, AO., Wilde, N., Gibson, M., Parks, A., Carrell, DT., Meikle, AW. (2008): Male obesity and alteration in sperm parameters. Fertil. Steril., 90: 2222-2225. |
| [5] | Magnusdottir EV, Thorsteinsson T, Thorsteinsdottir S, Maria Heimisdottir M, Olafsdottir K. (2005): Persistent organochlorines, sedentary occupation, obesity and human male subfertility. Hum. Reprod., 20 (1): 208-215. |
| [6] | Tsai, EC., Matsumoto, AM., Fujimoto, WY., Boyko, EJ. (2004): Association of bioavailable, free, and total testosterone with insulin resistance: influence of sex hormone-binding globulin and body fat. Diabetes Care, 27:861–868. |
| [7] | Pauli, EM., Legro, RS., Demers, LM., Kunselman, AR., Dodson, WC., Lee, PA. (2008): Diminished paternity and gonadal function with increasing obesity in men. Fertil. Steril., 90: 346-351. |
| [8] | Osuna, JA., Go´mez, PR., Arata, BG., Villaroe, lV., (2006): Relationship between BMI, total testosterone, sex hormone-binding-globulin, leptin, insulin and insulin resistance in obese men. Arch. Androl., 52: 355–361. |
| [9] | Sallmen, M., Sandler, DP., Hoppin, JA., Blair, A., Baird, DD. (2006): Reduced fertility among overweight and obese men. Epidemiology, 17: 520–523. |
| [10] | Nguyen, RH., Wilcox, AJ., Skjaerven, R., Baird, DD. (2007): Men’s body mass index and infertility. Hum Reprod, 22: 2488–2493. |
| [11] | Douchi, T., Kuwahata, R., Yamamoto, S., Oki, T., Yamasaki, H. and Nagata, Y. (2002): Relationship of upper obesity to menstrual disorders. Acta. Obstet. Gynecol. Scand., 81: 147–150. |
| [12] | Yura, S., Ogawa, Y., Sagawa, N., Masuzaki, H., Itoh, H. and Ebihara, K. (2000): Accelerated puberty and late-onset hypothalamic hypogonadism in female transgenic skinny mice overexpressing leptin. J. Clin. Invest., 105 (6): 749-755. |
| [13] | Sagae, S.C., Menezes, E.F .,. Bonfleur, M.L., Vanzela, E.C., Zacharias, P., Lubaczeuski, C., Franci,C.R. and Sanvitto, G.L. (2012): Early onset of obesity induces reproductive deficits in female rats. Physiology & Behavior, 105: 1104–1111. |
| [14] | Zhang, J.V., Ren, P.G., Avsian-Kretchmer, O., Luo, C.W., Rauch, R., Klein, C. and Hsueh, A.J. (2005): Obestatin, a peptide encoded by the ghrelin gene, opposes ghrelin's effects on food intake. Science, 310: 996-999. |
| [15] | Chanoine, JP., Wong, AC. and Barrios, V. (2006): Obestatin acylated and total ghrelin concentrations in the perinatal rat pancreas. Horm. Res., 66 (2): 81-88. |
| [16] | Dun, S.L., Brailoiu, G.C., Brailoiu, E., Yang, J., Chang, J.K. and Dun, N.J. (2006): Distribution and biological activity of obestatin in the rat. J. Endocrinol., 191: 481-489. |
| [17] | Pan, W., Tu, H. and Kastin, AJ. (2006): Differential BBB interactions of three ingestive peptides: obestatin, ghrelin, and adiponectin. Peptides, 27: 911–6. |
| [18] | Kaiya, H., Miyazato, M. and Kangawa, K. (2011): Recent advances in the phylogenetic study of ghrelin. Peptides, 32: 2155–2174. |
| [19] | Zhang, J.V., Jahr, H., Luo, C.W., Klein, C., Van Kolen, K., Ver Donck, L., De, A., Baart, E., Li, J., Moechars, D. and Hsueh, A.J. (2008): Obestatin induction of earlyresponse gene expression in gastrointestinal and adipose tissues and the mediatory role of G protein-coupled receptor, GPR39. Mol. Endocrinol., 22: 1464–1475. |
| [20] | Ren, A.J., Guo, Z.F, Wanga, Y.K., Lin, L., Zheng, X. and Yuan W.J. (2009): Obestatin, obesity and diabetes. Peptides, 30: 439–444. |
| [21] | Hassouna, R., Zizzari, P. and Tolle, V. (2010): The ghrelin/obestatin balance in the physiological and pathological control of growth hormone secretion, body composition and food intake. J. Neuroendocrinol., 22: 793–804. |
| [22] | Słupecka, M., Woliński, J., Herman, AP., Ochniewicz, P., Kornacka, MK. (2012): Biological role of obestatin in physiology and pathophysiology. Med. Wieku. Rozwoj., 16: 47–52. |
| [23] | Qi, X., Li, L., Yang, G., Liu, J., Li, K. and Tang, Y. (2007): Circulating obestatin levels in normal subjects and in patients with impaired glucose regulation and type 2 diabetes mellitus. Clin Endocrinol., 66 (4): 593-597. |
| [24] | Abou Fard, GM., Madi, NM. and Abo Zade, AA (2014): Circulating obestatin level in diabetic and obese rats. Tanta Medical Journal, 42 (1): 1–5. |
| [25] | Meszarosova, M., Sirotkin, AV., Grossmann, R., Darlak, K. and Valenzuela, K. (2008): The effect of obestatin on porcine ovarian granulosa cell. Anim. Reprod Sci., 108: 196-207. |
| [26] | Jahan, S., Sidrat, T., Ahmed, S., Wazir, H. and Ullah K. (2011): Effect of obestatin on morphometry of testes and testosterone secretion in male rats. African Journal of Biotechnology, 10 (39): 7717-7722. |
| [27] | Vigueras- Villasenor, R.M., Rojas-Castaneda S.J., Chavez-Saldana, M., Gutierrez-Pe, S., GarcıaCruz, M.R., Cuevas-Alpuche, O., Reyes-Romero, MM. and Zambrano, E. (2011): Alterations in the spermatic function generated by obesity in rats. Acta. Histochemica., 113: 214–220. |
| [28] | Freeman, M. E. (1988): The ovarian cycle of the rat. In: E. Knobil & J. Neil (eds.), Physiology of reproduction. Raven Press Ltd., pp. 1893-1928. |
| [29] | Marcondes, F.K., Bianchi, F.J., Tanno, A. P. (2002): determination of the estrous cycle phases of rats: some helpful considerations. Braz. J. Biol., 62 (4A): 609-614. |
| [30] | Long, J. A. and Evans, H. M. (1922): The estrous cycle in the rat and its associated phenomena. Memories of University of California, 6: 1-148. |
| [31] | Mandl, A. M. (1951):The phases of the oestrous cycle in the adult white rat. Journal of Experimental Biology, 28: 576-584. |
| [32] | Novelli, E., Diniz, Y., Galhardi, C. Ebaid, G., Rodrigues, H., Mani, F., Fernandes, A., Cicogna, A. and Novelli Filho, J. (2007): Anthropometrical parameters and markers of obesity in rats Laboratory Animals Ltd. Laboratory Animals, 41: 111–119. |
| [33] | Tietz, N.W. (1995): Clinical Guide to Laboratory Tests, 3rd Ed., W.B. Saunders Company, Philadelphia, PA 19106. |
| [34] | Starr J.I., Mako M.E., Juhn D. and Rubenstein A.H. (1978): Measurement of serum pro-insulin–like material: cross reactivity of porcine and human proinsulin. J. Lab. Clin. Med., 91: 691-692. |
| [35] | Matthews, D.R, Hosker, J.P., Rudensky, A.S., Naylor, B.A., Treacher, D.F. and Turner, R.C. (1985.: Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia, 28:412–419. |
| [36] | Flegg, H.M. (1973): An investigation of the determination of serum cholesterol by an enzymatic method. Ann. Clin. Biochem., 10:79-84. |
| [37] | Allain C., Poon L.S., Chan C.S., Richmond W. and Fu P.C. (1974): Enzymatic determination of total serum cholesterol. Clin. Chem., 20: 470-475. |
| [38] | Nagele U., Hagele E.O., Sauer G., Wiedmann E., Lehmann P., Wahlefeld A.W. and Gruber W. (1984): Reagent for the enzymatic determination of serum total triglycerides with improved lipolytic efficiency. J. Clin. Chem. Clin. Biochem., 22: 165-174. |
| [39] | Naito, H.K. (1989): Triglycerides in clinical chemistry: theory, analysis and correlation. Second edition by Kaplan LA and Pesce AJ, (U.S.A.), P. 997. |
| [40] | Warnick G.R., Benderson V. and Albers N. (1983): Selected methods. Clin. Chem., 10: 91-99. |
| [41] | Friedwald W.T., Levy R.I. and Fredrickson D.S. (1972): Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem., 18: 499-502. |
| [42] | Rebar, R.W., Morandini, I.C., Petze, J.E. and Erickson, G.F. (1982): Hormonal basis of reproductive defects in athymic mice: reduced gonadotropins and testosterone in males. Biol. Repro., 5: 1267-1276. |
| [43] | Casteilla L., Penicaud L. and Cousin B. (2001): Choosing an adipose tissue depot for sampling. Factors in selection and depot specificity. Methods. Mol. Biol., 155: 1–19. |
| [44] | Idris M.H, Budin S.B, Osman, M. and Mohamed, J. (2012): protective role of hibiscus sabdariffa calyx extract against streptozotocin induced sperm damage in diabetic rats. EXCLI. Journal., 11: 659-669. |
| [45] | Belsey M.A, Moshissi K.S, Eliasson R, Paulsen C.A, Callegos A.J. and Prasad M.R (1980): Laboratory manual for the examination of human semen and semen cervical mucus interaction. Press concern, 1-43. |
| [46] | Khaki A, Nouri M, Fathiazad F, Ahmadi-Ashtiani HR, Rastgar H and Rezazadeh (2009): Beneficial effects of quercetin on sperm parameters in streptozotocin-induced diabetic male rats. Phytother Res., 24 (9): 1285-1291. |
| [47] | Murthy, N.V., Wray, S.R., Melville, G.N., Wynter, H.H., Ram, N.V. and Haran, N.V. (1988): Testicular function in rats following immobilization stress. Int. J. Gynaecol. Obstet.., 22: 297-299. |
| [48] | Raghavendra, V., Tanga, F., Rutkowski, M.D., and DeLeo, J.A. (2003): Anti-hyperalgesic and morphine-sparing actions of propoentofylline following peripheral nerve injury in rats: mechanistic implications of spinal glia and proinflammatory cytokines. Pain, 104 (3): 655-664. |
| [49] | Wang, N., Luob, L.L., Xua, J.J., Xua, M.Y., Zhangb, X.M., Zhoub, X.L., Liu, W.J. and Fua, Y.C. (2014): Obesity accelerates ovarian follicle development and follicle loss in rats. metabolism clinical and experimental, 63: 94–103. |
| [50] | Ghanayem, B., Bai, R., Grace, E., Greg Travlos, K., and Hoffler, U. (2010): Diet-Induced obesity in male mice is associated with reduced fertility and potentiation of acrylamide-induced reproductive toxicity. Biology of reproduction, 82: 96–104. |
| [51] | Alvarez-Castroner P., Sangiao-Alvarellos S., Brandon-S. and Cordido F. (2011): Endocrine function in obesity. Endocrinol Nutr., 58 (8): 422-432. |
| [52] | Granata, R., Gallo, D., Luque, RM., Baragli, A., Scarlatti, F., Grande, C and Ghigo, E. (2012): Obestatin regulates adipocyte function and protects against diet-induced insulin resistance and inflammation. The FASEB Journal, 26: 3393–3411. |
| [53] | Guo, ZF., Zheng, X., Qin, YW., Hu, JQ., Chen, SP. and Zhang Z. (2007): Circulating preprandial ghrelin to obestatin ratio is increased in human obesity. J. Clin. Endocrinol. Metab; 92: 1875–1880. |
| [54] | Park, WH., Oh, YJ., Kim, GY., Kim, SE., Paik, KH., Han, SJ. (2007): Obestatin is not elevated or correlated with insulin in children with Prader-Willi syndrome. J. Clin. Endocrinol. Metab., 92 (1): 229-34. |
| [55] | Jarow, J.P and Zirkin, B.R. (2005):The androgen microenvironment of the human testis and hormonal control of spermatogenesis. Ann. N Y. Acad., 1061:208-20. |
| [56] | Yang, AJ., Cui, H., Cui, Y., Ye, HC. and Li Y. (2005): Effects on development of the testicle in diet- induced obesity rats. Wei. Sheng. Yan. Jiu., 34 (4): 477–479. |
| [57] | Wang, A.Y., Hickman, I.J., Richards, A.A., Whitehead, J.P., Prins, J.B. and Macdonald, G.A. (2005): High molecular weight adiponectin correlates with insulin sensitivity in patients with hepatitis C genotype 3, but not genotype 1 infection. Am. J. Gastroenterol., 100: 2717-2723.. |
| [58] | Erdemir, F., Atilgan, D., Markoc, F., Boztepe , O., Suha-Parlaktas, .B, Sahin, S.(2012): The effect of diet induced obesity on testicular tissue and serum oxidative stress parameters. Actas.Urol. Esp.,36(3):153-9. |
| [59] | Cabler, S., Agarwal, A., Flint, M. and du-Plessis, SS. (2010): Obesity: modern man’s fertility nemesis. Asian J Androl., 12: 1-10. |
| [60] | Oliva, A., Spira, A. and Multigner, L. (2001): Contribution of environmental factors to the risk of male infertility. Hum. Reprod., 16: 1768–1776. |
| [61] | Goyal, HO., Robateau, A., Braden, TD., Williams, CS., Srivastava, KK., Ali, K. (2003): Neonatal estrogen exposure of male rats alters reproductive functions at adulthood. Biol. Reprod., 68: 2081–2091. |
| [62] | Akingbemi, B.T. (2005): Estrogen regulation of testicular function. Reprod Biol Endocrinol., 3: 51. |
| [63] | Isidori, AM., Caprio, M., Strollo, F., Moretti, C., Frajese, G. and Isidori, A. (1999): Leptin and androgens in male obesity: evidence for leptin contribution to reduced androgen levels. J. Clin. Endocrinol. Metab., 84: 3673-380. |
| [64] | Tena-Sempere, M., Pinilla ,L., Zhang, FP., Gonzalez, LC., Huhtaniemi, I., Casanueva, FF., Dieguez, C. and Aguilar, E. (2001): Developmental and hormonal regulation of leptin receptor (Ob-R) messenger ribonucleic acid in rat testis. Biol. Reprod., 64: 634-643. |
| [65] | Tena-Sempere, M., Barreiro, M.L., Gonzalez, L.C, Gaytan, F., Zhang, F.P. and Caminos J.E. (2002): Novel expression and functional role of ghrelin in rat testis. Endocrinology, 143: 717–725. |
| [66] | Bucholtz, D.C., Chiesa, A., Pappano, W.N., Nagatani, S., Tsukamura, H., Maeda, K.I. and Foster, D.L. (2000): Regulation of pulsatile luteinizing hormone secretion by insulin in the diabetic male lamb. Biology of Reproduction, 62: 1248–1255. |
| [67] | Tanaka, T., Nagatani, S., Bucholtz, D.C., Ohkura, S., Tsukamura, H., Maeda, K. and Foster, D.L. (2000): Central action of insulin regulates pulsatile luteinizing hormone secretion in the diabetic sheep model. Biology of Reproduction, 62: 1256–1261. |
| [68] | Tanaka, M., Nakaya, S., Kumai, T., Watanabe, M., Matsumoto, N. and Kobayashi, S. (2001): Impaired testicular function in rats with diet-induced hypercholesterolemia and/or streptozotocin-induced diabetes mellitus. Endocrine Research, 27: 109–117. |
| [69] | Gromadzka-Ostrowska, J., Przepiorka, M. and Romanowicz, K. (2002): Influence of dietary fatty acids composition, level of dietary fat and feeding period on some parameters of androgen metabolism in male rats. Reproductive Biology, 2: 277–293. |
| [70] | Lu, Z.H., Mu, Y.M., Wang, B.A., Li, X.L., Lu, J.M., Li, J.Y., Pan, C.Y., Yanase, T. and Nawata, H. (2003): Saturated free fatty acids, palmitic acid and stearic acid, induce apoptosis by stimulation of ceramide generation in rat testicular Leydig cell. Biochemical and Biophysical Research Communications, 303: 1002–1007. |
| [71] | Vegiopoulos, A. and Herzig, S. (2007): Glucocorticoids, metabolism and metabolic diseases. Molecular and Cellular Endocrinology, 275: 43–61. |
| [72] | Cano, P., Jimenez-Ortega, V., Larrad, A., Reyes Toso, C.F., Cardinali, D.P. and Esquifino, A. (2008): Effect of a high-fat diet on 24-h pattern of circulating levels of prolactin, luteinizing hormone, testosterone, corticosterone, thyroid stimulating hormone and glucose, and pineal melatonin content, in rats. Endocrine, 33: 118–125. |
| [73] | Avelino-Cruz, JE., Flores, A., Cebada, J., Mellon, PL., Felix, R. and Monjaraz, E. (2009): Leptin increases L-type Ca2+ channel expression and GnRH-stimulated LH release in LbetaT2 gonadotropes. Molecular and Cellular Endocrinology, 298 (1–2): 57–65. |
| [74] | Brüning, JC., Gautam, D., Burks, DJ., Gillette, J., Schubert, M. and Orban, PC. (2000): Role of brain insulin receptor in control of body weight and reproduction. Science. 289: 2122–2125. |
| [75] | Xia, Y.X., Weiss, J.M., Polaack, S., Diedrich, K. and Ortmann, O. (2001): Interactions of insulin-like growth factor-I, insulin and estradiol with GnRH-stimulated luteinizing hormone release from female rat gonadotrophs. European Journal of Endocrinology, 144: 73–79. |
| [76] | Holdcraft, RW., and Braun, RE.( 2004): Hormonal regulation of spermatogenesis. International Journal of Andrology, 27: 335-342. |
| [77] | Garcia, M.C., Lopez, M., Alvarez, C.V., Casanueva, F., Tena-Sempere, M. and Dieguez, C. (2007): Role of ghrelin in reproduction. Reproduction, 133: 531-540. |
| [78] | Codoner-Franch, P., Tavrez-Alonso, S., Murria-Estal, R., Megas- Vericat, J., Tortajada-Girbés, M. and Alonso-Iglesias, E. (2011): Nitric oxide production is increased in severely obese children and related to markers of oxidative stress and inflammation. Atherosclerosis, 215: 475-480. |
| [79] | Farias, JG., Puebla, M., Acevedo, A., Tapia, PJ., Gutiérrez, E. and Zepeda, A. (2010): Oxidative stress in rat testis and epididymis under intermittent hypobaric hypoxia: protective role of ascorbate supplementation. J. Androl., 31: 314-321. |
| [80] | Brownlee M (2001): Biochemistry and molecular cell biology of diabetic complications. Nature, 414: 813-820. |
| [81] | Dohle, GR., Diemer, T., Giwercman, A., Jungwirth, A., Kopa, Z. and Krausz, C. (2012): EAU Guidelines on male infertility, 62: 324-332. |
| [82] | Galinier, A., Carriere, A., Fernandez, Y., Caspar-Bauguil, S., Periquet, B. and Periquet A. (2006): Site specific changes of redox metabolism in adipose tissue of obese Zucker rats. FEBS. Lett., 580: 6391–8. |
| [83] | Baccetti, B., La Marca, A., Piomboni, P., Capitani, S., Bruni, E., Petraglia, F. & De Leo, V. (2002): Insulin-dependent diabetes in men is associated with hypothalamo-pituitary derangement and with impairment in semen quality. Human Reproduction 17 2673–2677. |
| [84] | Abd El Samad A.A (2013): Role of aminoguanidine on the testis of streptozotocin-induced diabetic albino rat, a light and electron microscopic study. J. Histol., 33: 451 – 466. |
| [85] | Kasturi, S. S., J. Tannir, et al. (2008): The metabolic syndrome and male infertility. J. Androl., 29 (3): 251-259. |
| [86] | Yamamoto, D., Ikeshita, N., Daito, R., Herningtyas, EH., Toda, K. and Takahashi, K. (2007): Neither intravenous nor intracerebroventricular administration of obestatin affects the secretion of GH, PRL TSH and ACTH in rats. Regul Pept., 138 (2/3): 141–144. |
| [87] | Allwsh, T.A. and Mohammad, J.A. (2014): The effect of isolated obestatin hormone from plasma on some biochemical parameters in normal and diabetic rats. Raf. J. Sci., 25 (1): 82-100. |
| [88] | Aragno, M., Mastrocola, R.,Ghe, C., Arnoletti, E., Bassino, E., Alloatti, G. and Muccioli, M. (2012): Obestatin induced recovery of Myocardial dysfunction in type 1 diabetic rats: underlying Mechanisms. Cardiovascular Diabetology, 1: 129-13. |
| [89] | Raucci, R., Rusolo, F.;,Sharma, A., Colonna, G., Castello, G., Costantini, S. (2013): Functional and structural features of adipokine family. Cytokine, 61: 1-14. |
| [90] | Jerome, M., Goldman, A., Murr, S. and Ralph L. (2007): The Rodent Estrous Cycle: Characterization of Vaginal Cytology and Its Utility in Toxicological Studies. Birth Defects Research, 80: 84–97. |
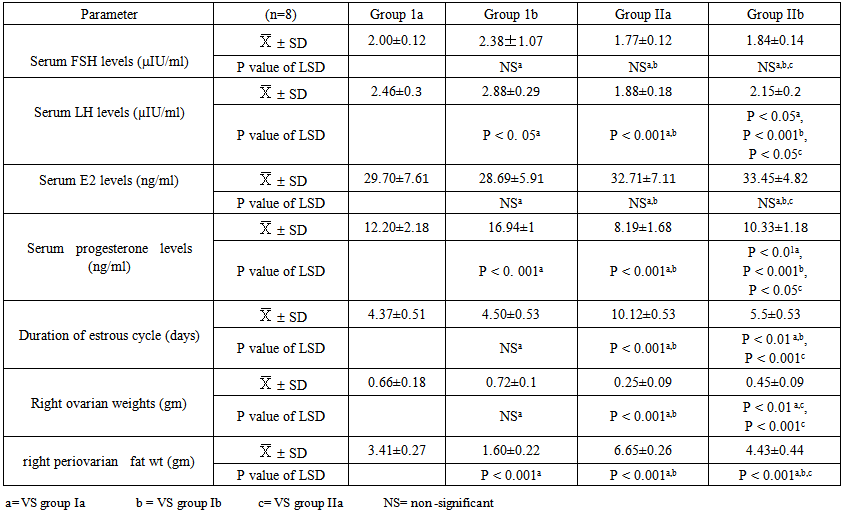
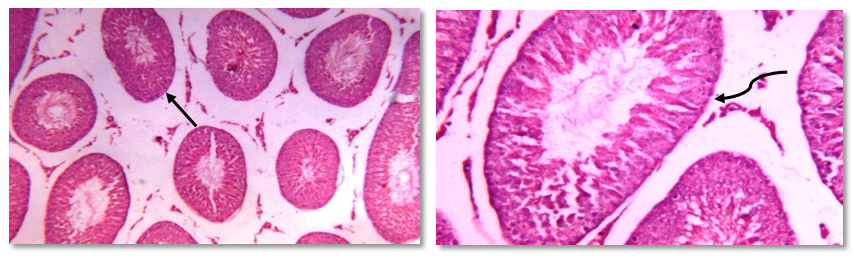
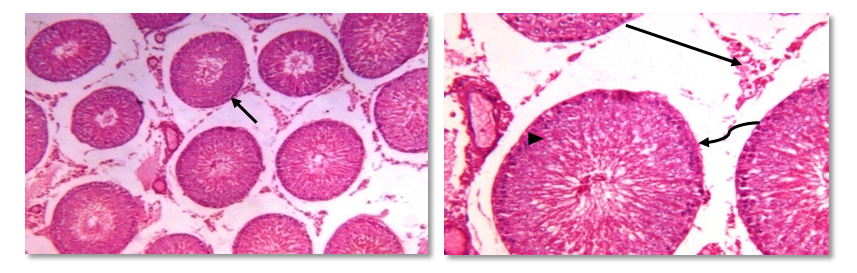
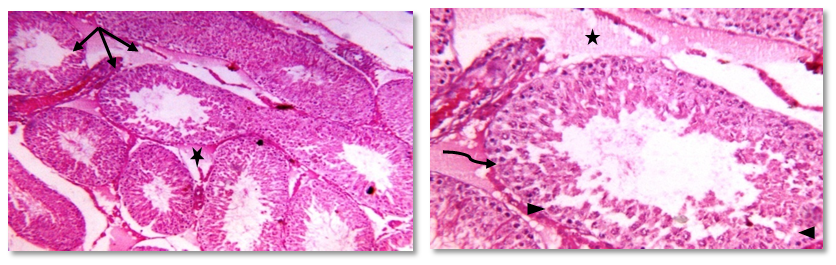
| [91] | Honnma H., Endo T., Kiya T., Shimizu A., Nagasawa K., Baba T., Fujimoto T., Henmi H., Kitajima Y., Manase K., Ishioka S., Ito E. and Saito T. (2010): Remarkable features of ovarian morphology and reproductive hormones in insulin-resistant zucker fatty (fa/fa) rats. Reprod. Biol. Endocrinol., 8: 73. |
| [92] | Poretsky, L., Cataldo, NA., Rosenwaks, Z. and Giudice, LC. (1999): The insulin related ovarian regulatory system in health and disease. Endocrine Reviews, 20: 535–582. |
| [93] | Brannian, JD., Furman, GM. and Diggins, M. (2005): Declining fertility in the lethal yellow mouse is related to progressive hyperleptinemia and leptin resistance. Reprod. Nutr. Dev., 45 (2): 143–50. |
| [94] | Lin, Q., Poon S.L., Chen, J., Cheng, L., HoYuen, B. and Leung, P.C. (2009): Leptin interferes with 3',5'-Cyclic Adenosine Monophosphate (cAMP) signaling to inhibit steroidogenesis in human granulosa cells. Reproductive Biology and Endocrinology, 7: 115. |
| [95] | Duggal, P.S., Van Der Hoek, K.H., Milner, C.R., Ryan, N.K., Armstrong, D.T., Magoffin, D.A. and Norman, R.J. (2000): The in vivo and in vitro effects of exogenous leptin on ovulation in the rat. Endocrinology, 141: 1971–1976. |
| [96] | Spicer, LJ., Chamberlain, CS. and Francisco, CC. (2000): Ovarian action of leptin: effects on insulin-like growth factor-I-stimulated function of granulosa and thecal cells. Endocrine, 12 (1): 53-59. |
| [97] | Balasubramanian, P., Jagannathan, L., Subramanian, M.,Ebony, T. Gilbreath, P.S., MohanKumar and Sheba M.J., MohanKumar, J. (2012): High fat diet affects reproductive functions in female diet induced obese and dietary resistant rats. Neuroendocrinol, 24 (5): 748–755. |
| [98] | Jones, S.M and Kazlauskas, A. (2001): Growth factor-dependent signaling and cell cycle progression. FEBS. Lett., 490: 110–116. |
| [99] | Naryzhny, S.N and Lee, H. (2001): Protein profiles of the Chinese hamster ovary cells in the resting and proliferating stages. Electrophoresis, 22: 1764–1775. |
| [100] | Muphy, B.D., Gevry, N., Ruiz-Cortes, T., Cote, F., Downey, B.R. and Sirois, J. (2001): Formation and early development of the corpus luteum in pigs. Reproduction, 58: 47–63. |
| [101] | Maga, G., Hubscher, U. (2003): Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J. Cell Sci., 116: 3051–3060. |
| [102] | Lagaud, GJ., Young, A., Acena, A., Morton, MF., Barrett, TD. and Shankley, NP. (2007): Obestatin reduces food intake and suppresses body weight gain in rodents. Biochem. Biophys. Res. Commun., 357 (1): 264–269. |
| [103] | Samson, WK., White, MM., Price, C. and Ferguson, AV. (2007): Obestatin acts in brain to inhibit thirst. Am. J. Physiol. Regul. Integr. Comp. Physiol., 292 (1): R637–R643. |
| [104] | Mony, A. and Batmanabane, M. (2013): Effect of obestatin on body weight, serum glucose and insulin levels in albino rats. Eur. J. Anat., 17 (2): 59-62. |
| [105] | Bascietto, C., Giannini, C., D’Adamo, E., de Giorgis, T., Chiarelli, F. and Mohn, A. (2011): Implications of gastrointestinal hormones in the pathogenesis of obesity in prepubertal children. J. Pediatr. Endocrinol. Metab., 25: 255–260. |
| [106] | Chen, CY., Doong, ML., Li, CP., Liaw, WJ., Lee, HF. and Chang, FY. (2010): SDA novel simultaneous measurement method to assess the influence of intracerebroventricular obestatin on colonic motility and secretion in conscious rats. Peptides, 31: 1113–1117. |
| [107] | Granata, R., Settanni, F., Gallo, D., Trovato, L., Biancone, L., Cantaluppi, V., Nano, R., Annunziata, M., Campiglia, P., Arnoletti, E., Ghe, C., Volante, M., Papotti, M., Muccioli, G. and Ghigo, E. (2008): Obestatin promotes survival of pancreatic beta cells and human islets and induces expression of genes involved in the regulation of beta-cell mass and function. Diabetes, 57: 967-79. |
| [108] | Lippl, F., Erdmann, J., Lichter, N., Tholl, S., Wagenpfeil, S., Adam, O. and Schusdziarra, V. (2008): Relation of plasma obestatin levels to BMI, gender, age and insulin. Horm. Metab. Res., 40: 806–812. |
| [109] | Gandhi, G.R., Stalin, A., Balakrishna, K., Ignacimuthu, S., Paulraj, M.G. and Vishal, R. (2013): Insulin sensitization via agonism of PPARγ and glucose uptake through translocation and activation of GLUT4 in PI3K / P-Akt signaling pathway by embelin in type 2 Diabetic rats. Biochimica. Biophysica. Acta., 1830: 2243-2255. |
| [110] | Manning, B. D., and Cantley, L. C. (2007): AKT/PKB signaling: navigating downstream. Cell, 129: 1261–1274. |
| [111] | Fogarty, S., and Hardie, D. G. (2010): Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim. Biophys. Acta., 1804: 581–591. |
| [112] | Gaidhu, M. P., Anthony, N. M., Patel, P., Hawke, T. J., and Ceddia, R. B. (2010): Dysregulation of lipolysis and lipid metabolism in visceral and subcutaneous adipocytes by high-fat diet: role of ATGL, HSL, and AMPK. Am. J. Physiol. Cell. Physiol., 298: C961–C971. |
| [113] | Kadowaki, T., Yamauchi, T., Kubota, N., Hara, K., Ueki, K., and Tobe, K. (2006): Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Invest., 116: 1784–1792. |
| [114] | Kadowaki, T., and Yamauchi, T. (2005): Adiponectin and adiponectin receptors. Endocr., 26: 439–451. |
| [115] | Muller, G., Ertl, J., Gerl, M., and Preibisch, G. (1997): Leptin impairs metabolic actions of insulin in isolated rat adipocytes. J. Biol. Chem., 272: 10585–10593. |
| [116] | Woo, MN., Bok, SH., Lee, MK., Kim, HJ., Jeon, SM., Do, GM., Shin SK, Ha TY. and Choi, MS. (2008): Anti-obesity hypolipidemic effects of a proprietary herb fiber combination (S&S PWH) in rats fed high-fat diets. J. Med. Food., 11: 169-178. |
| [117] | Kamal, AA. and Mohamed, AN. (2009): Effect of carnitine and herbal mixture extract on obesity induced by high fat diet in rats. Diabet & Metab Synd., 1: 1-17. |
| [118] | Grundy, SM. (2004): Metabolic complications of obesity. Obesity, Metabolic Syndrome, and Cardiovascular Disease. J. Clin. Endo. & Metab., 89 (6): 2595–2600. |
| [119] | Chen, CY., Asakawa, A., Fujimiya, M., Lee, SD. and Inui, A. (2009): Ghrelin gene products and the regulation of food intake and gut motility. Pharmacological Reviews, 61 (4): 430-481. |
| [120] | Katsiki, N., Mikhailidis, DP., Gotzamani-Psarrakou, A., Yovos, JG. and Karamitsos, D. (2011): Effect of Various Treatments on Leptin, Adiponectin, Ghrelin an Neuropeptide Y in Patients With Type 2 Diabetes Mellitus. Expert Opinion on Therapeutic Targets, 15: 401-20. |
| [121] | Al–Hakeim H.K. and Ali M.M. (2012): Low ghrelin level is associated with poor control and bad prognosis parameters in obese diabetic patients. Journal of Diabetology, 1 (5): 1-10. |
| [122] | Nagaraj, S., Peddha, MS. and Manjappara, UV. (2008): Fragments of obestatin as modulators of feed intake, circulating lipids, and stored fat. Biochemical and Biophysical Research Communications, 366: 731–737. |
| [123] | Nagaraj, S., Peddha, MS. and Manjappara, UV. (2009): Fragment analogs as better mimics of obestatin. Regulatory Peptides, 158: 143–148. |
| [124] | Agnew, A.; Calderwood, D.; Chevallier, OP.; Greer, B.; Grieve, DJ. And Green, BD. (2011): Chronic treatment with a stable obestatin analogue significantly alters plasma triglyceride levels but fails to influence food intake; fluid intake; body weight; or body composition in rats. Peptides, 32: 755–762. |
| [125] | Bourron, O., Daval, M., Hainault, I., Hajduch, E., Servant, J.M., Gautier, J.F., Ferre, P. and Foufelle, F. (2010): Biguanides and thiazolidinediones Inhibit stimulated lipolysis in human adipocytes through activation of AMP-activated protein kinase. Diabetologia, 53: 768-778. |
| [126] | Hsu, C.H., Liao, Y.L., Lin, S.C., Chou, P. (2012): Adiponectin level Predicts HDL–Cholesterol level in type 2 diabetes. The Open Atherosclerosis and Thrombosis, 5: 1-5. |
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML