-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Virology and Molecular Biology
p-ISSN: 2163-2219 e-ISSN: 2163-2227
2026; 15(3): 67-73
doi:10.5923/j.ijvmb.20261503.02
Received: May 10, 2026; Accepted: Jun. 6, 2026; Published: Jun. 16, 2026

Antioxidant Potential and Polyphenolic Profile of Endophytic Fungi Aspergillus Sydowii Strain AV5L and Penicillium Verrucosum AV6L Isolated from Aloe Vera
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLDilaram Ruzieva1, Nilufar Vakhabova1, Toshxon Gulyamova1, Gulchexra Rasulova1, Uchkun Ishimov2, Liliya Abdulmyanova1
1Institute of Microbiology, Academy of Sciences of the Republic of Uzbekistan, Tashkent
2Institute of Bioorganic Chemistry, Academy of Sciences of the Republic of Uzbekistan, Tashkent
Correspondence to: Nilufar Vakhabova, Institute of Microbiology, Academy of Sciences of the Republic of Uzbekistan, Tashkent.
| Email: |  |
Copyright © 2026 The Author(s). Published by Scientific & Academic Publishing.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Endophytic fungi are an important and underexplored source of multifunctional bioactive metabolites. In this study, we investigated the antioxidant potential of ethyl acetate extracts from Aspergillus sydowii AV5L and Penicillium verrucosum AV6L isolated from Aloe vera. The extracts exhibited strong concentration-dependent antioxidant activity in phosphomolybdenum, DPPH, and hydrogen peroxide scavenging assays. Notably, P. verrucosum AV6L demonstrated superior activity, showing phosphomolybdenum antioxidant capacity comparable to ascorbic acid (88.3 ± 2.3 AAE/mL at 1000 µg/mL) and exceeding it in hydrogen peroxide scavenging (78% at 1000 µg/mL). High levels of phenolic compounds and flavonoids were detected, and LC–MS/MS analysis revealed a diverse profile of polyphenols, including compounds with known antioxidant and urease inhibitory activity. These results highlight the potential of Aloe vera-associated endophytic fungi as a source of dual-function bioactive metabolites targeting both oxidative stress and enzyme-mediated pathogenicity.
Keywords: Endophytes, Urease, Metabolites, Antioxidant activity, Phenols, Flavonoids
Cite this paper: Dilaram Ruzieva, Nilufar Vakhabova, Toshxon Gulyamova, Gulchexra Rasulova, Uchkun Ishimov, Liliya Abdulmyanova, Antioxidant Potential and Polyphenolic Profile of Endophytic Fungi Aspergillus Sydowii Strain AV5L and Penicillium Verrucosum AV6L Isolated from Aloe Vera, International Journal of Virology and Molecular Biology, Vol. 15 No. 3, 2026, pp. 67-73. doi: 10.5923/j.ijvmb.20261503.02.
Article Outline
1. Introduction
- Endophytic microorganisms are increasingly recognized as an important source of biologically active secondary metabolites with potential relevance in pharmaceutical and biomedical research [1-3]. These microorganisms inhabit plant tissues without causing apparent harm and often establish complex symbiotic relationships with their host plants. As a result of co-evolution, endophytes can produce structurally diverse metabolites, including compounds similar to those synthesized by their host, as well as unique bioactive molecules that may possess notable pharmacological properties [4,5].Among medicinal plants, Aloe vera has attracted considerable attention due to its reported antioxidant, anti-inflammatory, antimicrobial, and gastroprotective properties [6,7]. These biological activities have been associated with the presence of phenolic compounds, flavonoids, and polysaccharides, which may contribute to reactive oxygen species (ROS) scavenging and modulation of inflammatory signaling pathways [8,9]. Endophytic microorganisms associated with Aloe vera may serve as alternative, sustainable sources of such compounds, as well as of potentially novel metabolites with diverse biological activities [10-11].One of the important molecular targets associated with infectious and inflammatory diseases is the enzyme urease. Urease is a nickel-dependent metalloenzyme that catalyses the hydrolysis of urea into ammonia and carbon dioxide [12-14]. This reaction contributes to the pathogenicity of several urease-producing microorganisms, including Helicobacter pylori infection, Proteus spp., and Yersinia enterocolitica [18-20]. Ammonia production may result in local pH elevation, epithelial damage, disruption of mucosal integrity, and activation of immune responses, including increased generation of reactive oxygen species.Thus, urease functions not only as metabolic enzyme but also as an important virulence factor associated with chronic inflammation, tissue damage, and increased risk of carcinogenesis. In the case of H. pylori infection, urease activity facilitates bacterial survival under acidic conditions while contributing to oxidative stress and activation of pro-inflammatory signalling pathways, including NF-κB. Infection with H. pylori stimulates innate immune responses, leading to the generation of reactive oxygen species (ROS) such as superoxide anion (O2−), hydrogen peroxide (H2O2), hydroxyl radicals (˙OH), and hypochlorous acid (HOCl). Increased ROS production in gastric cells and phagocytes following H. pylori exposure has been reported [22-23]. Elevated ROS levels in the gastric mucosa of infected patients may contribute to oxidative damage of proteins, DNA and lipids.Notably, many naturally occurring urease inhibitors belong to phenolic compounds, flavonoids, and related polyphenols [24]. These molecules may inhibit urease activity through interactions with Ni²⁺ ions located in the enzyme active site while also exhibiting antioxidant and anti-inflammatory properties. Therefore, metabolites possessing both urease inhibitory and antioxidant activities may represent promising candidates for reducing enzymatic virulence and oxidative stress simultaneously.Endophytic fungi associated with Aloe vera may represent a valuable source of multifunctional metabolites. In our previous studies, ethyl acetate extracts of Aspergillus sydowii strain AV5L and Penicillium verrucosum AV6L exhibited notable urease inhibitory activity, reaching up to 88,1% [25] inhibition. However, the antioxidant properties of these bioactive extracts have not yet been comprehensively investigated.Therefore, the present study aimed to evaluate the antioxidant activity of these endophytic fungal extracts using multiple in vitro assays, and to establish a possible relationship between their chemical composition and biological activity. A comprehensive approach combining antioxidant assays and LC–MS/MS analysis was applied to elucidate the bioactive potential of secondary metabolites produced by these endophytes.
2. Materials and Methods
2.1. Fungal Strains and Cultivation
- Endophytic fungi Aspergillus sydowii strain AV5L and Penicillium verrucosum AV6L were isolated from healthy leaves of Aloe vera collected in Tashkent, Uzbekistan. The isolates were previously identified by molecular methods and deposited in the NCBI GenBank database under accession numbers strain PQ726933 and strain PV065897 respectively.The endophytic fungi were cultivated under submerged fermentation conditions in 500 mL flasks containing 250 mL of potato dextrose broth and incubated for 7 days at 28°C on a rotary shaker.
2.2. Extraction of Secondary Metabolites
- 5 g of fungal biomass of endophyte was homogenized and transferred to a conical flask containing 50 ml of ethyl acetate, and left for 48 hours on the rotary shaker at room temperature. The mixture was filtered through filter paper (Whatman #1), after which the extract was dried on a rotary evaporator and redissolved in 1 ml of ethyl acetate. The resulting crude extract was used as a stock solution and stored at 4°C until further use.
2.3. Determination of Total Antioxidant Capacity
2.3.1. Phosphomolybdenum Assay
- Total antioxidant capacity (TAC) was evaluated by phosphomolybdenum assay according to Prieto et al (1999) [26], based on the reduction of Mo (VI) to Mo (V) and the formation of green phosphate/Mo(V) complex., with the refence to updated methodological overviews (Gulcin 2020) [27] with minor modifications. Briefly, 0.1 mL of each extract was mixed with 1 mL of reagent solution containing 0.6 M sulfuric acid, 28 mM sodium phosphate, and 4 mM ammonium molybdate. The reaction mixtures were incubated at 95°C for 90 min. After cooling to room temperature, absorbance was measured at 695 nm against a blank using a UV–Vis spectrophotometer (UV-5100, China). Results were expressed as ascorbic acid equivalents (AAE). All experiments were performed in triplicate.
2.3.2. DPPH Radical Scavenging Assay
- The free radical scavenging activity of the extracts was determined using the DPPH assay according to Blois [28]. Briefly, 1 mL of extract solution at different concentrations (100–1000 µg/mL) was mixed with 1 mL of 0.2 mM DPPH solution in ethanol. The reaction mixtures were incubated in the dark at room temperature for 30 min. Absorbance was measured at 517 nm. Ascorbic acid was used as a standard. The percentage of radical scavenging activity was calculated using the following equation: Scavenging activity (%) = (Ac − As) / Ac × 100, where Ac is the absorbance of the control and As is the absorbance of the sample.
2.3.3. Hydrogen Peroxide Scavenging Assay
- Hydrogen peroxide (H2O2) scavenging activity was determined according to the method of Ruch et al. [29] based on the ability of antioxidants to neutralize H2O2. A 40 mM hydrogen peroxide solution was prepared in phosphate buffer (50 mM, pH 7.4). Extract solutions (100–1000 µg/mL) were added to the H₂O₂ solution, and absorbance was measured at 230 nm after 10 min. The percentage of scavenging activity was calculated as: H₂O₂ scavenging (%) = [(Ai − At) / Ai] × 100, where Ai is the absorbance of the control, and At is the absorbance of the sample.
2.4. IC₅₀ Values Determination
- IC₅₀ values (the concentration required to inhibit 50% of radical activity) were calculated using nonlinear regression analysis based on concentration–response curves.
2.5. LC–MS/MS Analysis
- Chemical profiling of the extracts was performed using reversed-phase nano-liquid chromatography coupled with a Q-TOF mass spectrometer (Agilent 1200 LC system with CHIP-Q-TOF 6520B, Agilent Technologies). Separation was carried out on a Zorbax SB-C18 chip column (5 µm, 75 µm × 43 mm). The mobile phases consisted of: (A) 0.1% formic acid in 5% acetonitrile and (B) acetonitrile with 0.1% formic acid and 10% water. A gradient elution program (0–60% B) was applied. The flow rate was 0.6 µL/min. All experiment were carried out in triplicate.
2.6. Statistical Analyses
- Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Turkey’s post hoc test. Data were expressed as mean ±standard deviation (SD). Differences were considerate statistically significant at p < 0.05.
3. Results
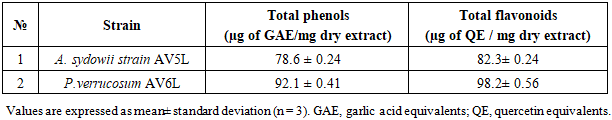
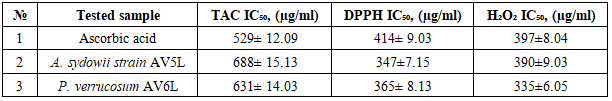
- Compounds combining urease inhibitory and antioxidant activities may provide a dual therapeutic effect by simultaneously suppressing enzymatic virulence and oxidative stress. In essence, having antioxidant activity ensures that the inhibitor not only works on the urease enzyme itself but also alleviates the oxidative damage that the pathogen has already inflicted on the host [30-32].The presented study was focused on the evaluation of the antioxidant activity of ethyl acetate extracts of Aspergillus sydowii strain AV5L and Penicillium verrucosum AV6L secondary metabolites, whose urease inhibitory activity reached 88%. The antioxidant activity was assessed through three different assays (TAC, DHPP, H2O2).The total antioxidant capacity assessed by the phosphomolybdenum method revealed a clear concentration-dependent increase in reducing power for both extracts. At the highest concentration (1000 µg/mL), P. verrucosum AV6L exhibited antioxidant activity (88.3 ± 2.3 AAE/mL) comparable to that of ascorbic acid (87.9 ± 2.4 AAE/mL), while A. sydowii strain AV5L showed slightly lower activity (76.0 ± 2.5 AAE/mL) (Figure 1).
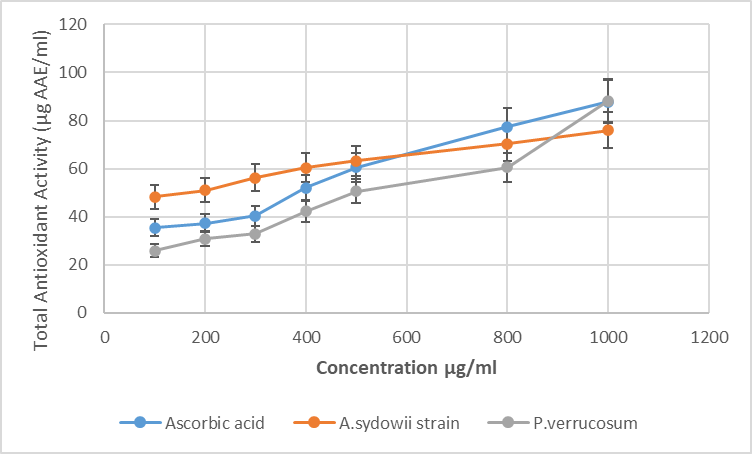 | Figure 1. The total antioxidant capacity of A. sydowii strain AV5L and P. verrucosum AV6L by the phosphomolybdate assay (n=3) |
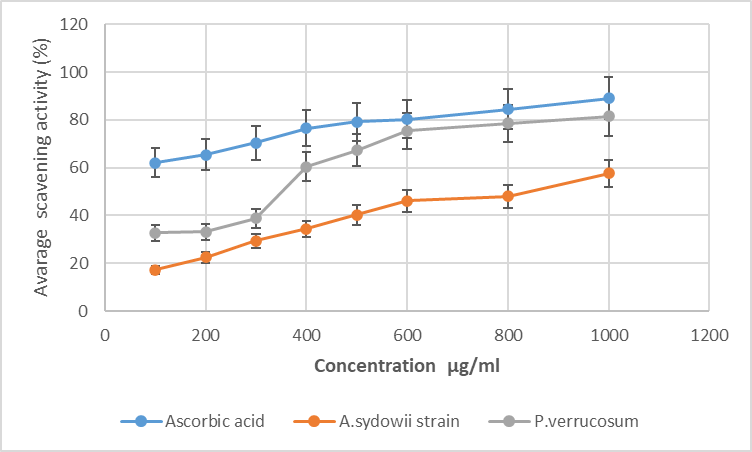 | Figure 2. Free radical scavenging activity of endophytic fungi A. sydowii strain AV5L and P. verrucosum AV6L п measured by the DPPH assay (n=3) |
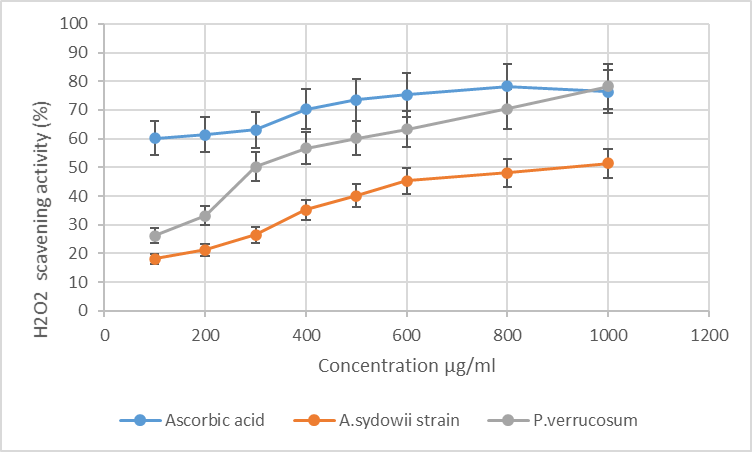 | Figure 3. Hydrogen peroxide (H₂O₂) radical scavenging activity of A. sydowii strain AV5L and P. verrucosum AV6L (n=3) |
|
|
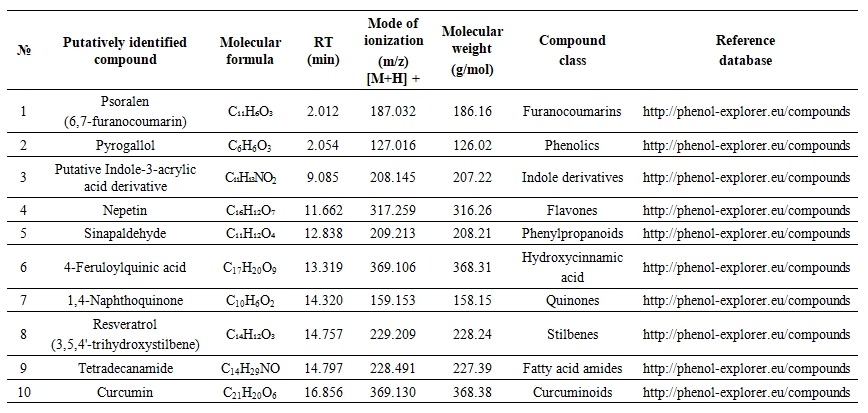 | Table 3. Mass Spectrometry Analysis of Endophytic Fungi Extracts A. sydowii Strain AV5L |
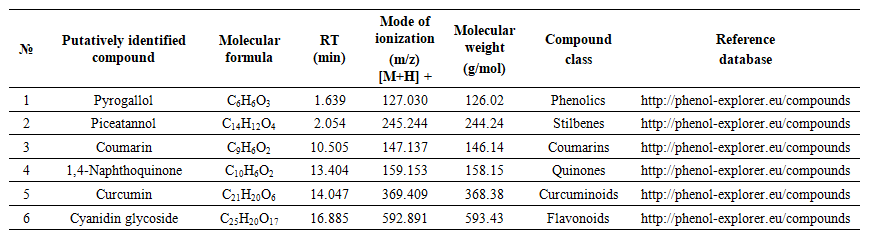 | Table 4. Mass Spectrometry Analysis of Endophytic Fungi Extracts Penicillium verrucosum AV6L |
4. Discussion
- The results obtained in this study clearly demonstrate that ethyl acetate extracts of the endophytic fungi Aspergillus sydowii strain AV5L and Penicillium verrucosum AV6L possess pronounced antioxidant activity, which is concentration-dependent and assay-specific. Among the investigated strains, P. verrucosum AV6L consistently exhibited superior antioxidant potential across all applied methods, particularly in hydrogen peroxide scavenging and DPPH assays.The observed antioxidant activity is strongly associated with the high content of phenolic compounds and flavonoids detected in the extracts. Numerous studies have demonstrated that phenolic compounds play a key role in neutralizing reactive oxygen species through electron donation and radical stabilization mechanisms (Shahidi & Ambigaipalan, 2015; Rice-Evans et al., 1997) [33,34]. In the present study, the higher total phenolic and flavonoid content in P. verrucosum AV6L directly correlates with its enhanced antioxidant capacity, supporting the widely reported relationship between polyphenol content and radical scavenging activity.Similar trends have been reported for endophytic fungi isolated from medicinal plants. For example, endophytes from Calotropis procera demonstrated a strong positive correlation between total phenolic content and antioxidant activity, including DPPH scavenging capacity and enzyme inhibition (Khiralla et al., 2015) [35]. These findings are consistent with our results and further confirm that phenolic-rich endophytic extracts represent a promising source of natural antioxidants.An important aspect of this study is the demonstrated link between antioxidant activity and urease inhibition. It is well established that many phenolic compounds, including flavonoids and stilbenes, exhibit dual biological activity, functioning both as antioxidants and enzyme inhibitors (Al-Rooqi et al., 2023) [36]. These compounds can chelate metal ions in the active site of urease while simultaneously reducing oxidative stress, which plays a critical role in the pathogenesis of infections caused by urease-producing microorganisms such as Helicobacter pylori.In this context, the dual functionality of the studied extracts is of particular interest. The combination of urease inhibitory and antioxidant activities suggests a synergistic therapeutic potential, where both enzymatic virulence and oxidative damage are targeted simultaneously. This concept has been previously described for plant-derived phenolics, but remains relatively underexplored for endophytic fungi.LC–MS/MS analysis revealed the presence of a diverse spectrum of putatively identified polyphenolic compounds, including resveratrol, flavonoids, phenolic acids, and quinones, which are widely recognized for their antioxidant and enzyme-inhibitory properties. Compound annotation was primarily based on LC-MS/MS fragmentation patterns and database comparisons. Compouds such as resveratrol and curcumin have been extensively reported to exhibit both antioxidant and urease inhibitory activity, further supporting the potential multifunctional nature of the detected metabolites (Bhat et al., 2021; Sharma et al., 2021) [37,38]. Overall, the results suggest that the observed biological activity of the studied endophytes may be associated with the synergistic interactions among multiple secondary metabolites rather than a single dominant compound. This synergistic effect is a well-known characteristic of natural extracts and often contributes to enhanced biological activity compared to isolated constituents.
5. Conclusions
- The present study demonstrates that endophytic fungi associated with Aloe vera, particularly Penicillium verrucosum AV6L, represent a promising source of biologically active secondary metabolites with significant antioxidant potential. The extracts exhibited strong concentration dependent radical- scavenging activity, reducing power, and hydrogen peroxide neutralization capacity.The observed biological activity is associated with the relatively high content of phenolic compounds and flavonoids, as indicated by quantitative analysis and LC–MS/MS profiling. The detected polyphenolic metabolites, some of with have been previously reported to exhibit antioxidant and urease inhibitory activities may contribute to the overall bioactivity of the extracts.Importantly, the combined antioxidant and urease inhibitory effects suggest that these endophytic fungi could be a dual-activity bioactive compounds acting on both oxidative stress enzyme-mediated pathogenic processes. However, further studies, including isolation of individual compounds and confirmation using authentic reference standards are required substantiate these activities and clarify their mechanisms on action. Further studies are needed to isolate and characterized individual compounds, clarify their mechanisms of action, and evaluate their biological effects in vivo. The present findings indicate that endophytic fungi isolated from Aloe vera, particularly Penicillium verrucosum AV6L, represent a potential source of biologically active secondary metabolites with significant antioxidant potential. The extracts demonstrated concentration-dependent radical-scavenging activity, hydrogen peroxide-neutralizing capacity, and reducing power, with some activities comparable to those of ascorbic acid under the tested condition.
ACKNOWLEDGEMENTS
- The authors acknowledge the support of the institute of Bioorganic Chemistry (Tashkent, Uzbekistan) for providing facilities for LC-MS analyses.