-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Virology and Molecular Biology
p-ISSN: 2163-2219 e-ISSN: 2163-2227
2026; 15(1): 1-6
doi:10.5923/j.ijvmb.20261501.01
Received: Dec. 23, 2025; Accepted: Jan. 16, 2026; Published: Jan. 22, 2026

Morphological and Molecular-Genetic Analysis of Some Species of Algaflora in Salted Soils of the Fergana Valley
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLSheraliyeva Dildora Nodir kizi1, Tokhtaboyeva Yulduzkhon Abdusattorovna2
1Phd Student, Namangan State University, Namangan, Uzbekistan
2Doctor of Biological Sciences, Associate Professor, Kimyo International University in Tashkent Branch Namangan, Namangan, Uzbekistan
Copyright © 2026 The Author(s). Published by Scientific & Academic Publishing.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

As salinity levels continue to rise worldwide, this poses a serious threat to agriculture. Such areas, including those in Uzbekistan, cover a significant amount of land. Studying the algal flora of these areas and utilizing its significance for agricultural purposes is one of the pressing tasks. In this regard, our research was conducted on cotton fields growing on saline soils in the Papsky and Mingbuloksky districts of the Namangan region of the Fergana Valley in Uzbekistan. Using molecular genetic analysis, we were able to identify five species of microalgae and cyanobacteria in samples collected from all regions. These species are adapted to saline soils and have been recorded as indicator species in cotton fields.
Keywords: Cyanobacteria, Microalgae, Diversity, ITS2, Soils, Salinity
Cite this paper: Sheraliyeva Dildora Nodir kizi, Tokhtaboyeva Yulduzkhon Abdusattorovna, Morphological and Molecular-Genetic Analysis of Some Species of Algaflora in Salted Soils of the Fergana Valley, International Journal of Virology and Molecular Biology, Vol. 15 No. 1, 2026, pp. 1-6. doi: 10.5923/j.ijvmb.20261501.01.
Article Outline
1. Introduction
- In Uzbekistan, the classification of saline soils includes types such as salt marshes and salt flats, which differ in terms of the depth of the salt deposits and their composition. Saline soils have a high concentration of water-soluble salts in the upper layer, whereas in saline soils, salts are located deeper and are present in the soil particle complex, especially in the sodium-rich horizon [FAO/ORG.2018]. Saline soils contain mineral salts in quantities that are harmful to plants. Crop growth begins to be inhibited when the salt content in the soil profile exceeds 0.25% of the soil mass [15].Saline soils cover a vast area of Uzbekistan, accounting for a significant portion of the land fund in such important agricultural regions as cotton-growing areas. Salinization, which occurs on irrigated land, has various causes, but regardless of its origin, salinization always has a negative impact on plant growth and development and on the properties of the soil itself. It destroys soil structure, impairs water-physical, physicochemical, and biological properties, affects microbiological activity and other properties, thereby causing soil degradation and plant death [15].In recent years, in many cotton-growing regions, particularly in Central Fergana, the soil reclamation and ecological condition of irrigated soils has deteriorated sharply, the level of mineralized groundwater has risen above the “critical” depth, processes of salinization and desertification have intensified, and the content of organic matter (humus) and plant nutrients, as well as the fertility and productivity of irrigated lands, have declined [15].
2. Materials and Methods of Research
- Description of the study areas. The Fergana Valley is an intermontane basin in the mountains of Central Asia, approximately 300 km long and up to 170 km wide, divided between three countries: Uzbekistan, Tajikistan, and Kyrgyzstan. The northern part of the Fergana Valley is home to the Namangan region of the Republic of Uzbekistan, which has a continental climate (dry summers and mild, wet winters). The average temperature in January is +4 °C, and in July +35 °C. Precipitation in the plains ranges from 135 mm to 370 mm per year, and in the foothills from 460 mm to 630 mm (Tukhtaboeva et al. 2025). Two areas within the region were studied: Papsky and Mingbuloksky, and five soil-algal samples were taken from cotton fields growing on saline soils. Below are the characteristics of the sampling points:1. Papskie Polya (Papski District). Altitude above sea level: 650–660 m. The soils are medium loamy typical gray-zems, washed to varying degrees and covered with gravel in places. The air temperature on the day of sampling was 12°C with a humidity of 47%. Two mixed soil-algal samples were collected from two locations: 1 sample – 40°77'24.20'‘N 70°96'08.56’'E.Isolation and cultivation of microalgae and cyanobacteria strains. To isolate strains from soil, we used the cumulative culture method with subsequent sowing on BG-11 agar medium. Next, through repeated streaking and isolation of individual colonies using a Pasteur pipette, algologically pure cultures were obtained [1]. The strains were then cultivated on solid BG-11 nutrient medium with nitrogen (pH = 7.0; agar 1.4%) in a climate chamber under standard conditions (temperature 23–25°C, light 60–75 μmol photons m–2 s–1, photoperiod 12 h). The strains studied were deposited in the All-Russian Collection of Microorganisms (VKM) under numbers VKM Al-505, 514, 515, 517, and 518.Light microscopy. The morphology and life cycles of algologically pure cultures were studied using light microscopy with a Leica DM750 microscope (Germany). The results of the observations were documented with photographs taken using a Leica Flexacam C3 color digital camera (Germany). The observation period ranged from 1 to 12 weeks. During morphological identification of microalgae strains, diacritical features such as thallus organization type, cell shape and size, chloroplast number and type, pyrenoids presence, mucilage presence and thickness, reproduction method, etc. were taken into account. During morphological identification of cyanobacterial strains, diacritical features such as thallus organization type, trichome width and polarity, cross-septum lacing, presence and type of mucous membranes, cell color, size, and shape, apical cell differences, etc. were taken into account. Leica Application Suite X software was used for morphometric measurements. To compare sizes, 100 cells from each strain were measured. Selected articles were used for morphological and molecular genetic identification of the green microalgae strain [4,5,6,7,8,9], cyanobacteria [11]. This study is based on the algae system adopted in the international electronic database Algae Base [2].Total DNA extraction, amplification, purification, and sequencing of amplicons. Total DNA was extracted from the strain using the DNeasy Plant Mini Kit (Qiagen, USA) according to the manufacturer's protocol. A ready-made Screen Mix-HS (Eurogen, Russia) was used for amplification. The conditions and primers for amplification of ITS2 [3] and the 16S rRNA gene [12,13,14] are shown in Table 1.
|

3. Results and Discussion
- Both strains had Bracteacoccus-like morphology.The cells of strain VKM Al-505 were single, spherical, 3.8‒15.5 μm in diameter (Fig. 1A). The cell wall does not thicken noticeably with age. Young cells contain 2–4 chloroplasts, while mature cells contain numerous chloroplasts without pyrenoids. Reserve substances – orange lipid droplets. Asexual reproduction by the formation of autospores (from 4 pieces in the mother cell), zoosporogenesis was not observed in culture.The cells of strain VKM Al-514 were single or in aggregates, spherical, 5.3‒27 μm in diameter (Fig. 1B). The cell wall is quite strong, but does not thicken noticeably with age. Young cells contain 2–4 chloroplasts, while mature cells contain numerous chloroplasts without pyrenoids. Reserve substances – orange lipid droplets. Asexual reproduction by the formation of autospores (from 4 pieces in the mother cell). Autospores often mature inside the sporangium wall and, after its destruction, can form cell aggregates. Zoospore formation has not been observed in culture.
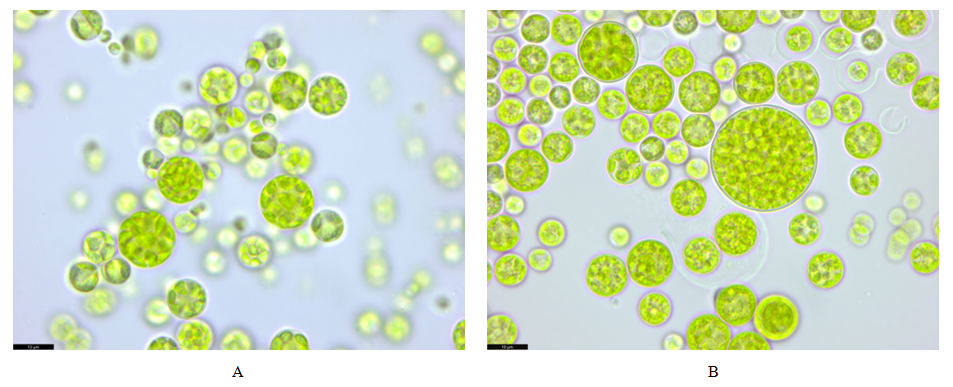 | Figure 1. Morphology of Bracteacoccus sp.VKM Al-505 (A) and 515 (B) strains. Scale bar: 10 μm |
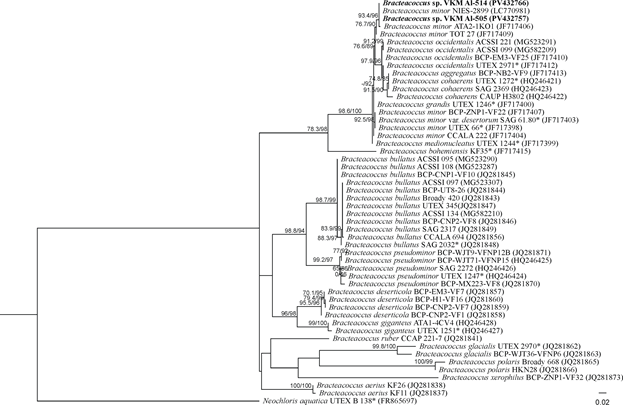 | Figure 2. Rooted phylogenetic tree of green microalgae of the genus Bracteacoccus, constructed using the ML method, based on the sequences of the internal transcribed spacer ITS2 (350 bp). SH-aLRT/UB values are given as statistical support for tree nodes. SH-aLRT and UB values less than 70% are not shown. Nucleotide substitution model: TIM2+F+G4. Symbols: * − authentic strains, studied strains are highlighted in bold |
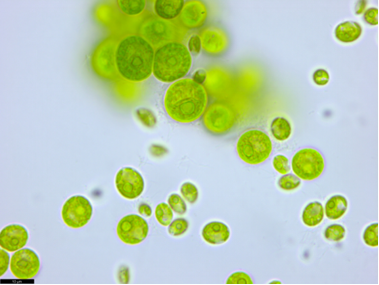 | Figure 3. Morphology of the Chlorococcum oleofaciens VKM Al-515 strain. Scale bar: 10 μm |
 | Figure 4. Rooted phylogenetic tree showing the taxonomic position of strain VKM Al-515, constructed using the ML method based on the sequences of the internal transcribed spacer ITS2 (274 bp). SH-aLRT/UB values are given as statistical support for tree nodes. SH-aLRT and UB values less than 70% are not shown. Nucleotide substitution model: TIM2e+G4. Designations: * − authentic strains, (T) – type species, the strain under study is highlighted in bold |
 | Figure 5. Morphology of Allocoleopsis franciscana strains VKM Al-517 (A) and 518 (B). Scale bar: 10 μm |
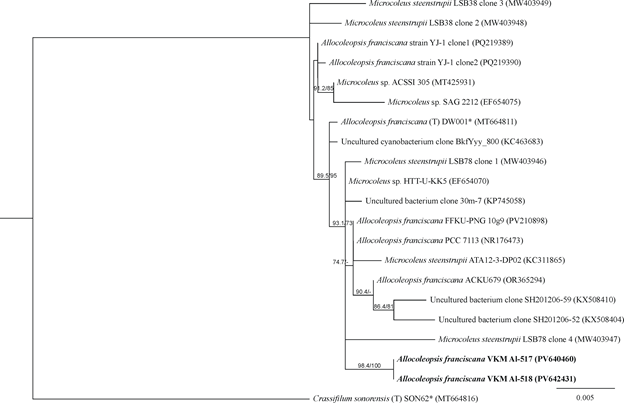 | Figure 6. Rooted phylogenetic tree of cyanobacteria of the genus Allocoleopsis, constructed using the ML method, based on 16S rRNA gene sequences (1493 bp). SH-aLRT/UB values are given as statistical support for tree nodes. SH-aLRT and UB values less than 70% are not shown. Nucleotide substitution model: HKY+F+I. Designations: * − authentic strains, (T) – type species, the studied strain is highlighted in bold |
4. Conclusions
- Thus, for the first time, morphological and molecular genetic analyses were used to study the diversity of microalgae cultivated on saline soils in the northern part of the Fergana Valley (Uzbekistan). Two strains of green microalgae (Chlorophyta) and one strain of cyanobacteria (Cyanobacteria) were detected. Only one species of microalgae and cyanobacteria has been identified within the species: Chlorococcum oleofaciens and Allocoleopsis franciscana. One other strain belongs only to the genus and requires further research: Bracteacoccus sp. The low species diversity of microalgae and cyanobacteria can be explained by both the high salinity and low fertility of the surrounding soils and the characteristics of the agrotechnical approach, which reveals only part of the true diversity of microorganisms. The research conducted may serve as a basis for the further development of highly functional consortia based on microalgae and cyanobacteria to improve and sustainably develop low-productivity, salinated, and degraded terrestrial ecosystems.