Zhigila A. D.1, Sawa F. B. J.2, Abdul S. D.2, Danailu G.2
1Botany Programme, Department of Biological Sciences, Gombe State University, Tudun Wada, Gombe, Gombe State, Nigeria
2Biological Sciences Programmes, Abubakar Tafawa Balewa University, Bauchi, Bauchi State, Nigeria
Correspondence to: Zhigila A. D., Botany Programme, Department of Biological Sciences, Gombe State University, Tudun Wada, Gombe, Gombe State, Nigeria.
| Email: |  |
Copyright © 2014 Scientific & Academic Publishing. All Rights Reserved.
Abstract
In the present study, an account was made to pollen morphology as an aid in taxonomy of twelve accessions of Sesamum indicum collected from Northern Nigeria. The accessions are ex-gombe 6, kenana 4, lale-duk, ex-gombe 5, ex-sudan, adawa-wula, adawa-ting (improved), adawa ting, ex-gombe 4, ex-gombe 1, ex-gombe 3 and ex-gombe 2; Keeping in view the importance and application of pollen study in taxonomy. Polleniferous materials were properly collected, identified and utilized for studies. The accessions show close relationship in their pollen surface characteristics having psilate sculpture. Pollen diameter ranges between 28.00µm in accession ex-gombe 4 to 16.75µm recorded by ex-sudan. Four types of pollen apertures were observed – periporate, inaperaturate, monoporate and dicolpate. Periporate recorded the highest frequency (81.82%) and (68.18%) in kenana 4 and lale-duk respectively, while monoporate recorded the lowest frequency (2.7%) in ex-gombe 6. Exine thickness ranged from 1.60µm to 0.95µm among the accessions. Kenana 4 appears to be the largest in exine thickness while ex-sudan is the smallest. These palynological features and the diversity were considered important characters to be used as an aid in taxonomy and other related disciplines of pure and applied sciences because this study provided some new dimensions towards further research in the discipline of palynology.
Keywords:
Sesamum indicum, Pollens, Morphology, Taxonomy, Accession
Cite this paper: Zhigila A. D., Sawa F. B. J., Abdul S. D., Danailu G., Diversity of Pollen Morphology in Accessions of Sesamum indicum L, International Journal of Modern Botany, Vol. 4 No. 1, 2014, pp. 22-28. doi: 10.5923/j.ijmb.20140401.03.
1. Introduction
Nigeria is one of the major producers of many oilseed crops like soybean, cotton, groundnut, mahogany, sesame seed, etc. Traditionally, Nigerians consume substantial quantity of edible oils mainly as a cooking medium. Among the oilseed crops, sesame has been cultivated for centuries, particularly in Asia and Africa, for its high content of edible oil and protein. It is commonly known as riɗɗi (Hausa), til (Hindi), hu ma (Chinese), sesame (French), goma (Japanese), gergelim (Portuguese) and ajonjoli (Spanish) Anilakumar et al (2010). Sesame, a member of Pedaliaceae family, is an annual shrub with white bell-shaped flowers with a hint of blue, red or yellow with or without branches (Martin and Leonard, 1967). It is grown for the production of seeds that are rich in oil content. It comes in a variety of colors, creamy-white to charcoal-black (Anilakumar et al., 2010). In general, the paler varieties of sesame seem to be more valued in West and Middle East, while the black varieties are prized in the Far East. Sesame is found in tropical, subtropical, and southern temperate areas of the world, particularly in India, China, South America and Africa. It has utmost economical importance and is primarily grown by small farmers in developing countries (FAOUN, 2012). The plant grows best in tropical climates, sandy, well drained soil with hot climate and moderate rainfall. It is propagated by seed sown in spring and takes about four months for the seeds to ripen fully. Sesame is a tropical herbaceous annual that grows 1-2 m tall. The plant has an unpleasant odour. The leaves vary from ovate to lanceolate and are hairy on both sides. The flowers are purple to whitish, resembling foxglove, followed by 3cm capsules/fruits containing numerous seeds (McCormick, 2001). Each plant may bear 15-20 fruits, which contain 70-100 seeds. It matures in 80–180 days when the stems are cut and hung upside down for the ripe seeds to fall out to be collected on mats.Pollen grains are reduced male gametophyte which upon pollination, produce pollen tubes that grow through the tissue of the pistil to effect fertilization and seed set (Muhammad et al., 2006). The production and dispersal of pollen have both biological and genetic implications for the quantity and genetic value of the seed produced. Hence pollen biology is of immense significance in plants improvement programmes as it determines gene flow and heterozygosity of the population, and those in turn determine genetic variability (Khanduri and Sharma, 2002). Pollen morphology has been used as the basis for phylogenetic studies in a number of angiosperm groups (Pandey and Misra, 2009). Pollen morphology can provide useful taxonomic characters if the surface morphology is sufficiently distinctive to allow reliable identification of taxa. It can also help to extend relationships on megamorphological grounds for species complexes at the family or generic level. The application of pollen morphology to plant systematic is comparatively a recent trend. It can provide a wealth of taxonomically useful information and would be useful in providing new information on infrageneric relationships. Bolick (1991) reported that the taxonomic and evolutionary importance of pollen morphology may be at specific, generic or higher level. The first palynological study in Pedaliaceae was that described by Erdtman (1952), who studied the pollen grains of 15 species belonging to 11 genera from this family. He showed that they are usually 5-13 colpate, oblate – subprolate, united in tetrads in Sesamothamnus and Sigmatosiphon species. Maria et al (1992) described the pollen morphology of Sesamothamnus lugardii. Ramakrishna and Bushan (2006) studied the pollen morphology of different genera including the genus Sesamum. Anjum and Qaiser (2002) reported that some of the pollen grains of the family Acanthaceae are similar to those of the family Pedaliaceae. In Nigeria, the pollen morphology of Pedaliaceae has not been investigated at length.The purpose of the present study is to provide detailed information on pollen morphology and to discover pollen morphological diversity in accessions of Sesamum indicum that would be helpful for taxonomists in particular and other fields of scientific research in general.
2. Materials and Methods
Twelve accessions of Sesamum indicum were collected from Northern Nigeria and planted. The list of voucher specimen was deposited in the GSUH. The accessions are ex-gombe 6, kenana 4, lale-duk, ex-gombe 5, ex-sudan, adawa-wula, adawa-ting (improved), adawa ting, ex-gombe 4, ex-gombe 1, ex-gombe 3 and ex-gombe 2; Keeping in view the importance and application of pollen study in taxonomy. Polleniferous materials were properly collected, identified and utilized for studies. The research work was conducted in the Biology Laboratory of the Biological Sciences Programme, School of Science, Abubakar Tafawa Balewa University, Bauchi. The study was confined to pollen morphology with taxonomic description of the accessions. Plant specimens were collected from the research farm of the aforementioned institution. For pollen morphological studies, pollens were separated from anthers and smeared on microscope slides. The preparations were stained with 1% safranin, immersed in a few drops of ethanol for 5 minutes to remove waxy substance from the pollens and 50% glycerol and observed under a light microscope. The fresh polleniferous materials used were according to a special technique known as Wodehouse technique (Ronald, 2002). Slides prepared according to this standard method are known as reference slides. Common adhesive that transparent finger nail polish was used to seal off the edges of reference slides. Permanent slides for pollen reference have been deposited in the Department of Biological Sciences, Abubakar Tafawa Balewa University, Bauchi, Nigeria. Different parameters studied under light microscope for pollen morphology were qualitative characters such as pollen type, shape, presence or absence of colpi, pores, spines, sculpturing and ornamentation. Quantitative characters include pollen polar diameter, equatorial diameter and exine thickness. Using 35 fields of view at x40 objective as quadrat, the numbers of pollens were noted to determine the frequency of the different pollen types present. Frequency of each pollen type was expressed as percentage occurrence of such pollen types based on all occurrences using this formula: P/y x 100 where p = the occurrence of each pollen type in the field of view; y = total occurrences of all pollen types. The density of pollen types was determined as the number of pollen types per square millimetre (mm2). Specimens were photographed under oil immersion using Am scope microscopic camera (MU1000, FMA050) and Kodak digital camera (Kodak Easy Share C913). Measurements were made using an ocular micrometer and all the measurements were taken in μm. Pollen description and identifications followed those of Willard et al (2004).
3. Results and Discussion
A summary of pollen morphological data in the accessions of sesame is presented in Table 1. Micrographs are presented in Figs 1 – 21. The accessions show close relationship in their pollen surface characteristics having psilate sculpture. Pollen size varies among these accessions very considerably. The pollen grains are consistently radially symmetrical, isopolar and isodiametric in equatorial view indicating a great variation in pollen size among the accessions and it was found that pollen shapes in polar view were prolate, spheroid, circular or semi-circular (Figs 1 – 21). The difference in size and shape between pollen grains found in this work were also determined in sunflower (Atlagic, 1990), rape seed (Atlagic et al., 2009) and sugar beet (Mezei et al., 2005). Canak and Parabucski (1966) stated that pollen grain size in most plant species varies in the range of 15 to 50 μm, and that the largest pollen grain is (150-200 μm) in gourd, which was confirmed by the measuring taken in this work. Four types of pollen apertures were observed – periporate, inaperaturate, monoporate and dicolpate. Ex-Gombe 4 has the highest pollen diameter (28.00µm) and ex-sudan can be distinguished due to the lowest (16.75µm) in equatorial diameter. Periporate recorded the highest frequency (81.82%) and (68.18%) in kenana 4 and lale-duk respectively, while monoporate recorded the lowest frequency (2.7%) in ex-gombe 6 (Table 1). There is a range of variation in exine thickness which has proved to be useful at specific level. Exine thickness ranged from 1.60µm to 0.95µm among the accessions. Kenana 4 appears to be the largest (1.60µm) in exine thickness while ex-sudan is the smallest (0.95µm). Pollens and spores are provided with a special wall or sporoderm Meo (2005). The outer part of this - the exine is usually resistant to decay, and is furnished with diverse processes, pits or net-like thickenings. Meo (2005) concluded that many forms of pollen grains, including variations in their shape, size, number and arrangement of apertures, and the fine sculpturing of their exine, are adaptations to help the pollen grain better perform its function of fertilizing the female gametophytes and forming seeds that will give rise to new generations of plants. Rodriguez (2000) studied the influence of genotype on pollen grain morphology in sesame. The results obtained by this author were very significant because they show that pollen grains from dehiscent anthers (free pollen) and non dehiscent anthers did not differ in shape or exine wall pattern but did differ by the dimensions of pollen. They also indicated the influence of environment on pollen dimensions. Plants can often be identified by means of these characteristics. It was found that pollen shape in polar view were prolate spheroid (adawa-ting (improved), adawa-ting, lale-duk and ex-gombe 1), circular (ex-sudan, ex-gombe 5, lale-duk, ex-gombe 6 and kenana 4) or semi-circular (lale-duk, ex-gombe 3, adawa-wula and kenana 4). Meo (2005) considered these palynological features as important characters to be used as an aid in taxonomy and other related disciplines of pure and applied sciences. Weber (1998) reported a systematic approach to categorising pollen types that enable the reader to recognise pollen characteristics of the most common botanical source. By applying a visual gestalt utilising grain size, shape, surface structure and internal detail, one is able to identify pollen source to appropriate botanical taxonomic level. It was found from this work that pollen morphology was specific for most of the analyzed accessions with slight overlapping of some similar characters; for instance, periporate and inaperaturate pollen apertures were observed in all the accessions. 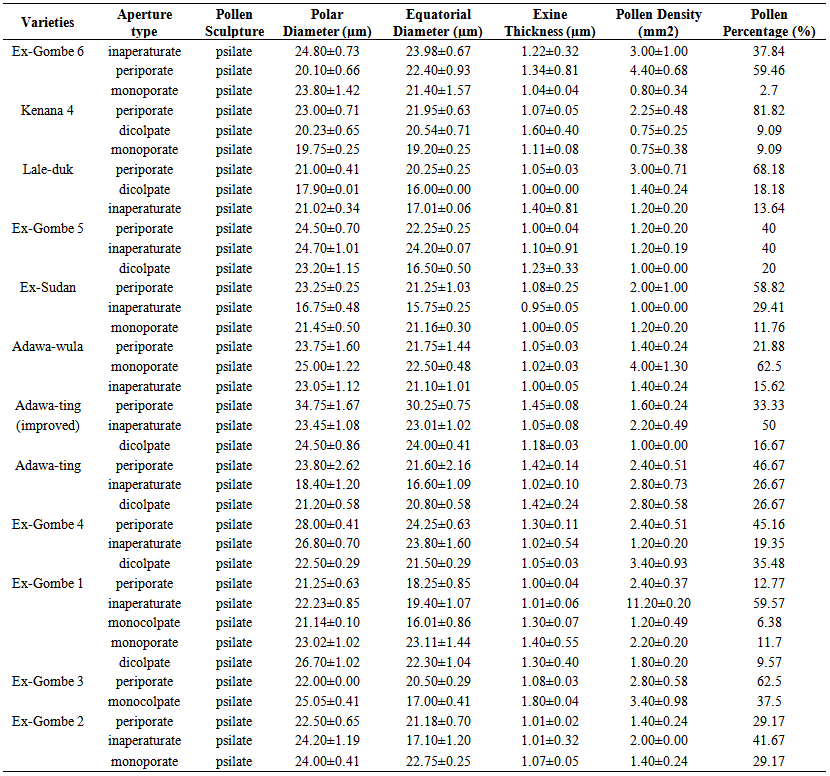 | Table 1. Pollen characteristics in accessions of S. indicum |
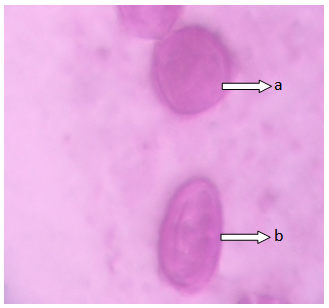 | Figure 1. Polar view of the pollen grain of accession ex-gombe 6 showing (a) circular (b) spheroid (x100) |
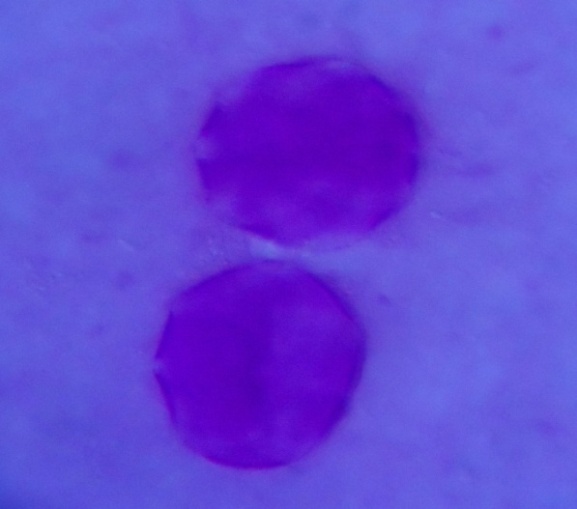 | Figure 2. Polar view of periporate pollen grains of accession kenana 4 (x100) |
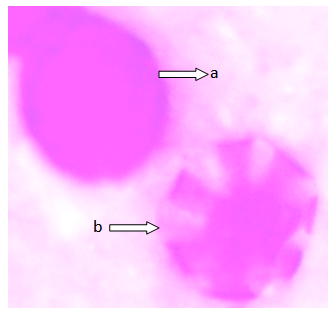 | Figure 3. Polar view of the pollen grain of accession ex-gombe 5 showing (a) inaperaturate (b) periporate (x100) |
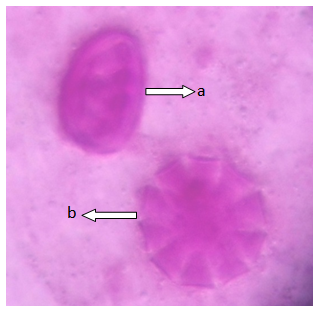
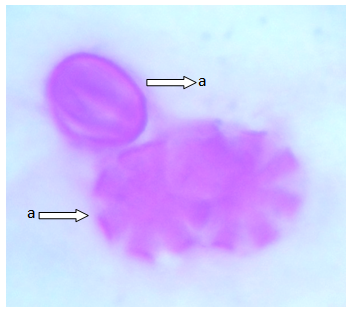 | Figure 4. Polar view of the pollen grain of accession lale-duk showing (a) dicolpate (b) periporate (x100) |
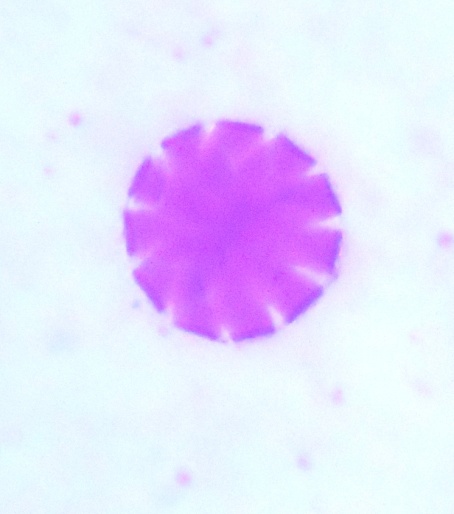 | Figure 5. Polar view of periporate pollen grains of accession ex-sudan (x100) |
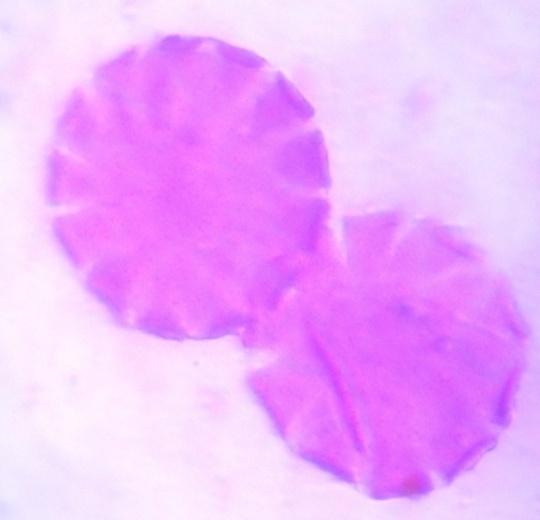 | Figure 6. Polar view of periporate pollen grains of accession adawa-wula (x100) |
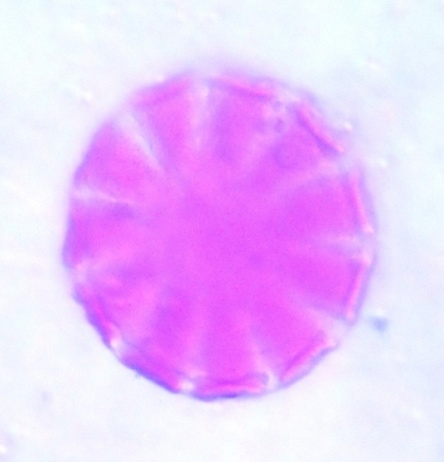 | Figure 7. Polar view of periporate pollen grains of accession adawa-ting (improved) (x100) |
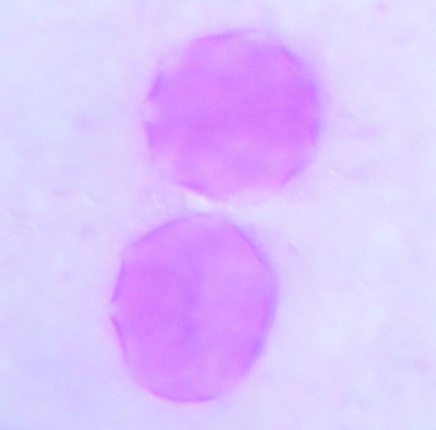 | Figure 8. Polar view of periporate pollen grains of accession adawa-ting (x100) |
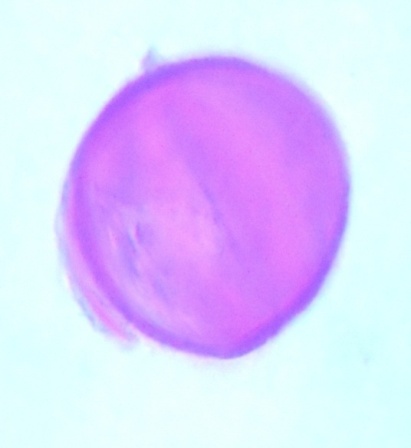 | Figure 9. Polar view of inaperaturate pollen grains of accession ex-gombe 4 (x100) |
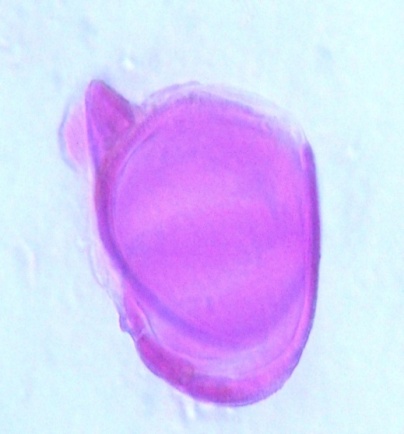 | Figure 10. Polar view of inaperaturate pollen grains of accession ex-gombe 1 (x100) |
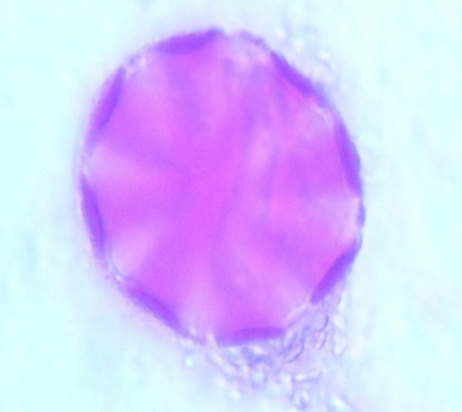 | Figure 11. Polar view of periporate pollen grains of accession ex-gombe 2 (x100) |
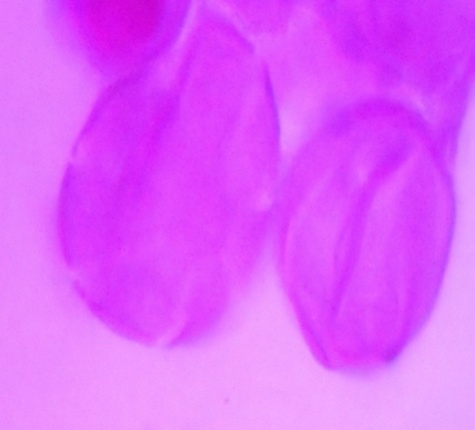 | Figure 12. Polar view of dicolpate pollen grains of accession adawa-ting (improved) (x100) |
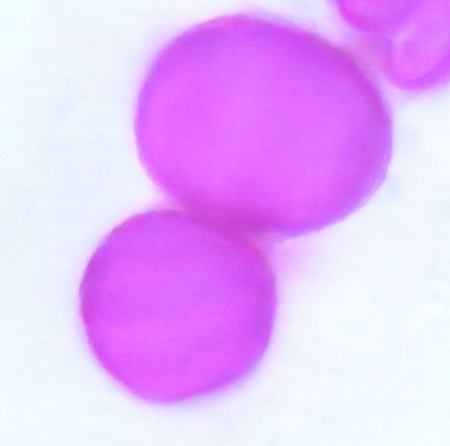 | Figure 13. Polar view of inaperaturate pollen grains of accession e-gombe 5 (x100) |
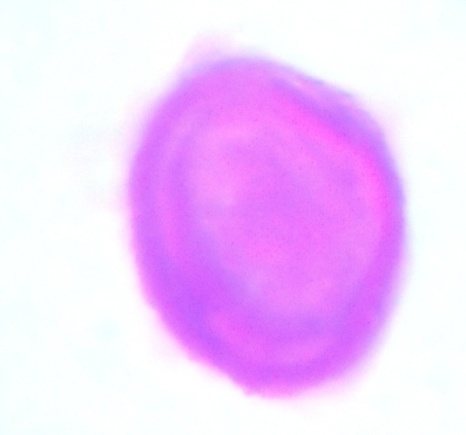 | Figure 14. Polar view of inaperaturate pollen grain of accession ex-gombe 3 (x100) |
 | Figure 15. Polar view of periporate pollen grains of accession ex-gombe 6 (x100) |
 | Figure 16. Polar view of periporate pollen grains of accession ex-gombe 1 (x100) |
 | Figure 17. Polar view of periporate pollen grains of accession e-gombe 5 (x100) |
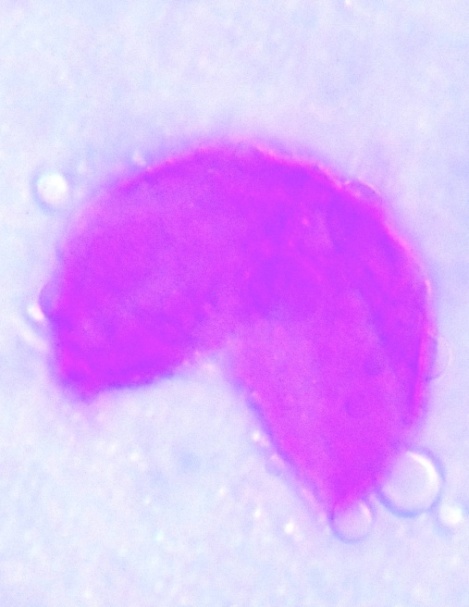 | Figure 18. Polar view of monoporate pollen grains with semi-circular shape of accession lale-duk (x100) |
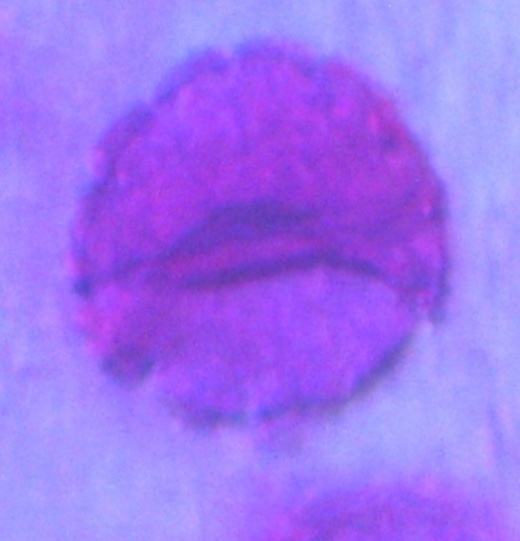 | Figure 19. Polar view of monocolpate pollen grains of accession ex-gombe 1 (x100) |
 | Figure 20. Polar view of (a) inaperaturate (b) periporate pollen grains of accession ex-gombe 3 (x100) |
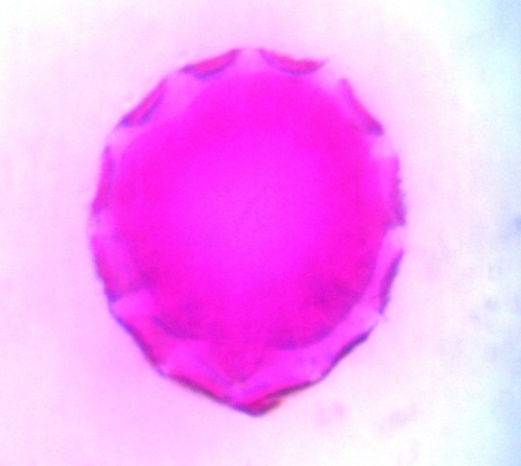 | Figure 21. Polar view of periporate pollen grains of accession lale-duk (x100) |
This information in combination with cross compatibility represents valuable and usable data for breeding programs. Atlagic et al (2009) suggested that studying the pollen grains morphology and cross compatibility of different genotypes (cultivars, populations) from the same species, as well as the cross compatibility of different species is necessary in further research of the oil species collection. These palynological features together with other taxonomic evidences were considered important characters to be used as an aid in taxonomy.
References
| [1] | Anilakumar, K.R., Pal, A., Khanum, F. and Bawa, A.S. (2010). Nutritional, medicinal and industrial uses of Sesame (Sesamum indicum L.) seeds - an overview. Biochemistry and Nutrition Discipline, Defence Food Research Laboratory, Mysore, India. |
| [2] | Anjum, P. and Qaiser, (2002). M.LXVII Acanthaceae, Journal of Botany, Special Issue 42:175-191. |
| [3] | Atlagic, J. (1990): Pollen fertility in some Helianthus L. species and their F1 hybrids with the cultivated sunflower. Helia, 13(13):47-54. |
| [4] | Atlagic, J., A. Marjanovic-Jeromela, R. Marinkovic, and S. Terzic (2009): Cytogenetic studies of cytoplasmatic male sterility in rapeseed. Proceedings of the 12th International Rapeseed Congress, March 26-30, Wuhan, China, Vol. I, 66-70. |
| [5] | Bolick, M. (1991). Pollen diameter, exine thickness and ultrastructure type in tribes of the Compositae. – Compositae News Letter, 19: 17-21. |
| [6] | Canak, M., and S. Parabucski (1966): Botanika. Naucna knjiga, Beograd, 1-407. |
| [7] | Erdtman, G. (1952) “Pollen morphology and plant taxonomy (Angiosperms).” Chrronica Botanica Co.,Waltham, Mass. .434-358. |
| [8] | Food and Agriculture Organization of the United Nations (FAOUN) (2012). "Food and Agricultural commodities production: Countries by commodity. |
| [9] | Khanduri, V.P. and Sharma, C.M. (2002). Pollen production, microsporangium dehiscence and pollen flow in Himalayan Cedar (Cedrus deodara Roxb. Ex D. Don). Annals of Botany, 89:587-593. |
| [10] | Maria, S., Juan, S. and Danielle, L. (1992). Plant Systematics and Evolution, 183:67-81. |
| [11] | Martin J. H. Leonard W. H. (1967). Miscellaneous industries crops. In: Principles of field crop production. Macmillan, New York. pp 922-924. |
| [12] | McCormick, H. (2001). Sesame Seeds (Sesamum indicum). Consumer products. www.mccormick.com/retail.nsf/4c144. |
| [13] | Meo, A. A. (2005). Palynological studies of selected genera of the tribes of Asteraceae from Pakistan. A thesis (Ph. D) submitted to the Quaid-i-Azam University, Islamabad – Pakistan. |
| [14] | Mezei, S., J. Atlagic, and L. Kovacev (2005): Pollen viability and meiosis in tetraploid populations of sugar beet (Beta vulgaris L.). Journal of Genetics & Breeding, 59(2):157-164. |
| [15] | Muhammad Z., Mir A.k., Mushtaq A. and Shazia S. (2006). Palynological and taxonomic studies weeds from Flora of Rawalpindi. Pakistan Journal of Weed Science Research, 12 (1-2): 99-109. |
| [16] | Ramakrishna, H. and Bushan, M. (2006). Geophytology, 1/2 53-59 LM. |
| [17] | Rodriguez, I.L. (2000): Correlations Between the Structure and Function of Pollen Grains of Four Species of Angiosperms. In: Scanning Electron Microscopy and X-ray Analysis. Edd. Briggs, D., Brady, J. Northampton, MA: Smith College, 2000. |
| [18] | Ronald, O.K. (2002). Pollen and spores. 2nd ed. American Association of Stratographic Palynologists, 13-21. |
| [19] | Weber, R. W. (1998). Pollen identification. Annal-Allegy, Asthma-Immunology, 80(20): 141 – 147. |
| [20] | Willard, D.A., Cooper, S.R., Gamez, D. and Jenshen, J. (2004). Atlas of pollen and spores of the Florida everglades. Palynology, 28: 175 – 227. |
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML