-
Paper Information
- Next Paper
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Ecosystem
p-ISSN: 2165-8889 e-ISSN: 2165-8919
2015; 5(2): 59-65
doi:10.5923/j.ije.20150502.03
Artemia Occurrence, Salinity and Ionic Rates in Saline Crater Lakes of Western Uganda
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLMujibu Nkambo1, Fred W. Bugenyi2, Sauda Nayiga3
1Aquaculture Research and Development Center - Kajjansi, National Fisheries Resources Research Institute (NaFIRRI), Kampala, Uganda
2Department of Biological Sciences, College of Natural Sciences (CONAS), Makerere University, Kampala, Uganda
3Faculty of Law Islamic University in Uganda, Mbale, Uganda
Correspondence to: Mujibu Nkambo, Aquaculture Research and Development Center - Kajjansi, National Fisheries Resources Research Institute (NaFIRRI), Kampala, Uganda.
| Email: |  |
Copyright © 2015 Scientific & Academic Publishing. All Rights Reserved.
Seven selected saline crater lakes in Western Uganda were surveyed for Artemia occurrence, salinity and ionic rates both in the wet and dry season. The Studied lakes included lakes Katwe, Munyanyange, Nyamunuka, Murumuri, Bunyampaka, Bagusa, and Kikorongo. In the field cyst-like materials along the lake shores and foam suspected to have Artemia cysts from the surface of each of the studied saline Crater Lake were collected in a plastic sample bottle. Presence of cysts was examined using the density separation technique. A zooplankton net of 50µm was used to filter water samples to be used for analysis of presence of Artemia biomass. Water samples for ionic composition analysis were taken from at least three geo-references points on each of the studied lakes. In-situ measurements of salinity were taken at each point of sample collection in each of the studied lakes. In the laboratory, collected water samples were analysed following APHA standard operating procedures. Seasonal differences in salinities and ionic rates within the studied lakes were tested for significance using a two tailed, paired t test. All the studied lakes had neither Artemia cysts nor biomass at the time of sampling both in the dry and wet season. Both in the dry and wet season, the salinity in all the study lakes was much higher than that of sea water while the ionic rates of Cl-/SO42- , Cl-/Na+ + K+, and Mg2+/Ca2+in all the study lakes were all less than that of sea water (table 1 & figure 2). The dry season salinity of all the study lakes was found to be significantly higher than the salinity in the wet season (t-value = 0.0205). There were no significant seasonal differences in the ionic rates of Cl-/SO42- , Cl-/Na+ + K+, and Mg2+/Ca2+in the studied lakes with t-values of 0.5665, 0.4082 and 0.4523 respectively. There is need for continuous monitoring of these saline lakes for the presence of Artemia resources since many other factors like temperature, the presence of wading migratory birds reported to be the dispersion agent for Artemia were witnessed in these lakes.
Keywords: Artemia Occurrence, Ionic rates, Saline lakes
Cite this paper: Mujibu Nkambo, Fred W. Bugenyi, Sauda Nayiga, Artemia Occurrence, Salinity and Ionic Rates in Saline Crater Lakes of Western Uganda, International Journal of Ecosystem, Vol. 5 No. 2, 2015, pp. 59-65. doi: 10.5923/j.ije.20150502.03.
Article Outline
1. Introduction
- Aquaculture production is one sure route suggested to bridge the gap between the ever increasing fish demand and the declining capture fisheries production. One of the requirements for sustainable aquaculture industry is reliable source of fish seed. Fish seed requires readily available source of live food especially at the larval stage and this has been one of the bottle necks for the development of Uganda’s aquaculture. Although Artemia has been reported to be of great importance as part of live food chain in culture of fish larvae [33], [31], [21], the high cost of imported Artemia cysts which ranges between US$50 to 100 per Kg [10], [21] has prohibited its wide use in Ugandan hatcheries hence the very low survival rates in the majority of Uganda’s hatcheries. Despite Uganda being endowed with hypersaline (salt lakes) environments which are among the reported natural habitats for Artemia [33] [8], [24], a comprehensive survey to explore the occurrence of local Artemia resources and possibility for Artemia production in these lakes is still lacking. The purpose of this study is therefore to explore the occurrence of local Artemia resources and ionic rates of these saline lakes systems as baseline information for commercial Artemia production in Uganda. This information on occurrence of local Artemia resources and ionic rates of these saline lakes is very useful in guiding the development of a commercial Artemia production industry in Uganda, which would offer a cheaper option for the currently very expensive imported Artemia resources. Past studies on these saline lakes in Western Uganda have focused on limnology [18], [20] and salt production in these lakes [13], [14]. Although there has been speculation about occurrence of Artemia in saline lakes of Uganda [21], Currently there is no grey literature on the occurrence of Artemia resources and ionic rates of these saline crater lakes in western Uganda. To guide this study, the following hypotheses were used; (1) There are no locally occurring Artemia resources in the saline crater lakes of Western Uganda. (2) Ionic rates of the saline crater lakes of western Uganda do not favor Artemia occurrence.Artemia distribution Artemia are primitive arthropod having segmented bodies with broad leaf-like appendages known as thoracopodes [10]. Artemia are commonly referred to as brine shrimps. They are micro-zooplankton belonging to anostracan crustaceans with a reported occurrence on all continents except in the Antarctica [33], [4], [22]. They are reported to naturally occur in salt lakes [30] and hypersaline environments with salinities as high as 340gl-1, a wide geographical distribution [21], [22], varying ionic compositions, climatic conditions and altitude [3]. The geographical distribution of Artemia was reported to be world-wide with the exception of the Antarctica [1] [3]; [22] and it is related to the different types of climate. World-wide over 600 Artemia sites are reported to have been discovered with more efforts on going to identify more sites [1] [3] [21]. Artemia is reported to have both sexually and parthenogenetically reproducing strains with a number of environmental parameters most notably salinity being reported to be responsible for the switch between giving live young ones and production of diapausing cysts [12]. It is the diapausing cysts that can withstand the harsh adverse conditions [1], stay viable for several years and are able to hatch into live nauplii on return of favorable conditions [12].Much as Artemia is reported to have a wide geographical distribution, very little is known about its distribution in Sub Saharan Africa [34]. Due to favorable climatic conditions most African countries are considered to have a huge potential for Artemia occurrence. Much of the information about Artemia distribution in Africa has been on Artemia franciscana, Artemia salina and the parthenogenetic strain in North African countries like Morocco, Egypt, Tunisia, Libya among others [3], [22]. Recently Artemia resources have also been reported in the east African region at the coast in Tanga, Tanzania and in Kenya with no concrete information on its occurrence in Uganda. Artemia importance Artemia as live food within the aquaculture industry dates back to the 1930s after it was discovered to be of high nutritional value to the new hatched fish larvae [5], [10]. Owing to its high nutritional value and convenience in use, Artemia is acknowledged world-wide as one of the most important starter feed within hatcheries of shell and fin fish. Despite the various efforts to find a suitable starter feed to replace Artemia because of cyst shortage and its high costs yielding varying levels of success [10], up to date no suitable artificial starter feed to replace Artemia has been found [21]. Currently over 2000 tons of Artemia dry cysts are reported to be used annually within the aquaculture industry world-wide [10] [21] and its demand is projected to increase with the ever increasing demand for fish leading to increased aquaculture production. It is this projected increment in demand for Artemia as a starter feed which has lead to numerous search for alternative starter feeds as well as new sites for Artemia production [10]. Through bio-encapsulation and enrichment, Artemia is currently being used as a medium to supply essential nutrients like vitamin, high unsaturated fatty acids [10], medication and antibiotics to shrimp and fish larvae within the aquaculture industry [12], [23], [25]. Besides its use in aquaculture industry, it has also been reported to improve salt quality because of its capacity to filter algae from the salt pans [26]. Ogello et al., (2014) reported salt production companies to have agreed that Artemia production is vital in controlling the algal blooms within the salt production pans. Having natural occurring Artemia resources would play a vital role in improve the salt quality in these saline lakes where commercial salt extraction is currently being carried out.Ionic ratesIonic composition and ionic rates are among the factors affecting salinity levels in these lakes and therefore affecting the occurrence of various organisms in these systems including fish, Artemia and others [6]. Having an understanding of the ionic composition and ionic rates in these saline lakes provides useful information which might explain the occurrence and absence of various organisms in these systems. These inland saline systems have been generally classified into carbonate and bicarbonate (CO32- + HCO3-, sulphate (SO42- and chloride (Cl-) depending on the dominant anion [18], [2] with Artemia population reported to occur in all these different categories [33], [28] with a reported preference for waters with chlorides as the dominant anions [7].
2. Materials and Methods
- Study AreaAll the studied Lakes Katwe (029.87033°E, 00.13217°S), Munyanyange (029.88591°E, 00.13513°S), Nyamunuka (029.98743°E, 00.09344°S), Bagusa (030.17958°E, 00.09793°S), Murumuri (029.99186°E, 00.07323°S), Bunyampaka (030.12819°E, 00.03765°S) and Kikorongo (030.01228°E, 00.01190°S) are found in western Uganda and belong to Katwe – Kikorongo volcanic field. With the exception of Kikorongo which is sometimes fresh, all the studied lakes are alkaline saline in nature [20], with a reported average depth ranging between <1 to 6m [14]. Artemia Cysts OccurrenceData collection in this study was carried out over a one year sampling period between the 26th of February, 2014 and 25th February, 2015. Since this area has been reported to receive a bimodal rainfall pattern [11], [13] during data collection the two wet seasons (March – May and September – December) and the two dry seasons (December-February and June-August) were considered. In the field cyst-like materials along the lake shores and foam suspected to have Artemia cysts from the surface of each of the studied saline Crater Lake were collected from at least four sites, put in plastic sample bottles before being carried to the laboratory for further analysis. In the laboratory at Kajjansi Aquaculture Research and Development Center (KARDC), cyst-like materials and foam suspected to contain Artemia cysts were tested for Artemia cysts using the density separation technique. Under the density separation technique, cyst-like materials and foam suspected to contain Artemia cysts were washed in a strong brine solution (above 45mgl-1), followed by washing in fresh water (0mgl-1) and finally washing in a strong brine solution (above 45mgl-1). Artemia cysts float in a strong brine solution, sink when washed in fresh water and float again when washed in a strong brine solution. Any cyst-like materials which floated in strong brine solution, sunk in fresh water solution and floated again on washing in strong brine solution were taken for further microscopic observation to be confirmed as Artemia cysts. Any collected material that behaved contrary to the above mentioned observation for cysts under the density separation technique were considered to have no Artemia cysts present.Artemia biomass occurrenceFour litres of surface water were collected and concentrated by filtering it through a 50µm zooplankton net. The plankton net sieve was rinsed with distilled water after filtering the four litres of the collected lake water and the rinsed water together with the residue collected in a plastic bottle. This was fixed by adding ethanol (90%) equal to a tenth of the total volume of the sample collected in the plastic sample collection bottle. In the laboratory at KARDC, the collected water samples were observed under a dissecting microscope for any Artemia biomass present.Salinity, Ionic composition and ratesSurface water samples for chemical composition analysis of each of the studied lakes were collected using a Van Dorn water sampler from three geo-referenced sampling points and stored in pre-rinsed Nalgene bottles for analysis in the laboratory. In the field all water samples collected in pre-rinsed Nalgene bottles were kept in a cooler box containing dry ice and later transferred to the National Fisheries Resource Research Institute (NaFIRRI) laboratory in Jinja. In-situ measurements of salinity were taken using hand-held refractors (0- 160mgl-1). In the laboratory the collected water samples were analysed for the following parameters: bicarbonate (HCO3), carbonate (CO3), Sulphate (SO4), Chloride (Cl-), Fluoride (F), Sodium (Na), Magnesium (Mg), Calcium (Ca), Potassium (K), and Sodium, Magnesium, Calcium, Potassium, and Chloride (Cl) following APHA 1975 standard Operating procedures. The measured salinities in the dry and wet season as well as the ionic rates were compared with those of sea water. Seasonal differences in salinities and ionic rates within the studied lakes were tested for significance using a two tailed, paired t test.
3. Results
- Artemia occurrence All the studied lakes had neither Artemia cysts nor biomass at the time of sampling in both the dry and wet season.Salinity and Ionic rates The Cl-/SO42- ionic rates in this present study ranged between 0.5±0.2and 1.73±0.7observed in lakes Bagusa and Murumuri in the dry season and these far less than the 7.16 of sea water.Both in the dry and wet season, the salinity in all the study lakes was much higher than that of sea water while the ionic rates of Cl-/SO42- , Cl-/Na+ + K+, and Mg2+/Ca2+ in all the study lakes were all less than that of sea water (table 1 & figure 2). The dry season salinity of all the study lakes was found to be significantly higher than the salinity in the wet season (t-value = 0.0205). Lakes Munyanyange and Nyamunuka were found to be dry and empty during the dry season sampling period while Lake Kikorongo was found to be fresh with salinity of 0mgl-1 during the wet season sampling (table 1 & figure 1). There were no significant seasonal differences in the ionic rates of Cl-/SO42- , Cl-/Na+ + K+, and Mg2+/Ca2+ in the studied lakes with t-values of 0.5665, 0.4082 and 0.4523 respectively.
|
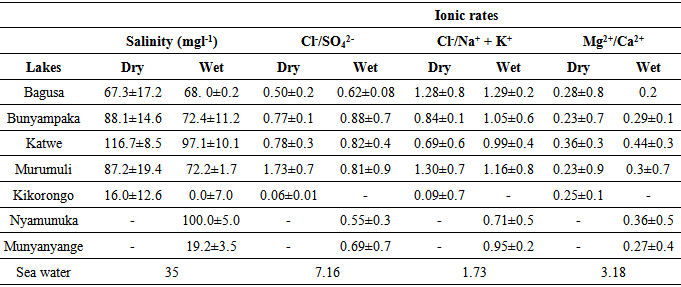
 | Figure 1. Dry and wet season salinity changes in the studied saline lakes |
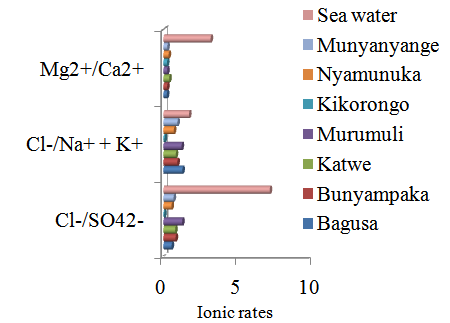 | Figure 2. Ionic rates of the different studied lakes in comparison with sea water |
4. Discussion
- Artemia occurrence This study constitutes the very first comprehensive survey of the occurrence of local Artemia resources in the saline lakes of western Uganda. Despite the presence of the flamingos which are among the report dispersion agents for brine shrimp eggs [8] and the speculation of occurrence of Artemia resources in these lakes [21], none of the study lakes was found to have Artemia biomass or cysts at the time of sampling both during the dry and wet season. Wind and wading migratory birds have been reported as the main vectors for the spread and maintenance of gene flow between distant Artemia populations [12], [1], [28]. The presence of migratory birds such as flamingos in these lakes gives an anticipation of Artemia being inoculated in these lakes by these birds. Predation, lack of appropriate food, habitat activities and other abiotic factors might be responsible factors responsible for the absence of Artemia in these studied lakes. In a study of the distribution and historical biogeography of Artemia lech, 1819 in Ukraine, adults and larvae of some Coleoptera (Hygrotus ennegrammus (Ahrens, 1833), Hemiptera (Coricidae), Ostracods Eucypris inflata (Sars 1903) and other invertebrates were reported to have probably eaten away all the Artemia in the lakes with salinities below 110mgl-1 [28]. In the same way zooplanktons, other invertebrates and birds most notably the lesser flamingos (Phoeniconaias minor) which live in these saline crater lakes might be eating away all the Artemia inoculated in these lakes. Although Artemia has been reported to exist at salinities higher than 340mgl-1 [28], the very high salinities might not be the only limiting factor. Other abiotic factors like dissolved oxygen might also be limiting. Dissolved oxygen is reported to significantly decrease with increasing salinity [28] and this could be another reason to explain the absence of Artemia in these hypersaline lakes in this current study. Planktons serve as the primary source of food for different groups of aquatic animals [29], with their biodiversity reported to decrease with increasing salinity [17], [15]. Although some bacteria strains have been reported as potential diets for Artemia [35], phytoplanktons are reported to be the main constituent of natural Artemia diets [26]. Because these currently studied lakes had very high salinities, it is expected that the planktonic biodiversity in these lakes is very low hence the lack of appropriate food organism for Artemia in these lakes. This further supported by findings from a study on feeding biology of parthenogentic Artemia in shallow seasonally stratified hypersaline lake in Western Australia where limited planktonic primary productivity was reported [27]. Although salts are part of integral biochemistry of life in aquatic environments, at very high concentration salt is reported to have toxic effects on these organisms [9]. Artemia being non selective filter feeders [27], [10], [35], it would require too much energy to filter large volumes of water so as to capture the very few existing phytoplanktons. Much as habitat activities have been reported to play a role in creating new habitats for Artemia, anthropogenic activities such as mining have also been reported to have destructive effect on the old exiting habitats [28]. This very explanation might be extended to explain the absence of Artemia in lakes like Katwe and Bunyampaka where salt mining constitutes to be one of the commercial activities in these lakes.Salinity, ionic composition and rates The significantly high salinity levels in the dry season incomparision to the wet season could be explained the fact that evaporation rates are higher in the dry season while precipitation become less in the dry season which leads to evaporative concentration of the salts in these lakes. Evaporative concentration lead to In this present study it was found that ionic rates of these saline systems are different from those of sea water. This is supported by Alcocer and Escober (1993) who reported the majority inland saline systems to have different ionic composition from that of sea water. Ionic composition in these inland systems has been reported to be largely determined by the incoming waters from water inflow and rainfall, dissolved material from the soils, rocks, and sediment of the drainage and lake basin [6]. Climate, geography, and topography of the drainage basin are some of the other factors reported to affect ionic composition in these saline waters [2], [32]. Worldwide ionic composition and ionic rates of saline waters have been reported to vary but with regional consistence [6]. This regional consistence is witnessed in this present study where all the studied lakes had Cl-/SO42- rates less than 2, Cl-/Na+ + K+ less than 1.5, and Mg2+/Ca2+ less than 0.5 both in the dry and wet sampling season (table 1). In a study of fluoride concentration and mineralogical composition of East African Magadi (trona), Lakes Magadi in Kenya, and Natron in Tanzania were found to have very similar mineral composition [19]. This further emphasizes the regional consistence in ionic composition in these saline water bodies. From the ionic rates in this current study it indicates that sea water has more chloride ions than any of the current studied lakes but this cannot be fronted to explain the absence of Artemia in this present study since Artemia resources have been reported in hypersaline environments [3] other than the sea and these might be of varying ionic rates from the of sea water. The ability of Artemia occurrence in environments of diverse ionic concentrations is well documented and this one of the factors reported to be responsible for diversity in phenotypes within the Artemia strains [3]. Never the less it should be noted that the majority of the study lakes have been reported to be carbonate - sulphate dominated in terms of anion composition [20], yet a number of literature have reported the brine shrimps to have a preference of carbonate- chloride dominated waters [7]. This might be pointed to as an explanation for the absence of Artemia resources in these currently studied lakes.
5. Conclusions
- No Artemia resources were found in this present study. The salinity and ionic rates of the lakes considered under this study were found to be different from that of sea water. There is need for continuous monitoring of these saline lakes for the presence of Artemia resources since many other factors like temperature, the presence of wading migratory birds reported to be the dispersion agent for Artemia were witnessed in these lakes.
ACKNOWLEDGEMENTS
- Special thanks and acknowledge is reserved for the National Agricultural Research Organisation (NARO), National Fisheries Resources Research Institute (NaFIRRI), Kajjansi Aquaculture Research and Development Center (KARDC), Department of Biological Science, College of Natural Sciences (CONAS) of Makerere University, Uganda Wildlife Authority (UWA) and Katwe-Kabatoro Community for the various support offered while carrying out this research.