-
Paper Information
- Previous Paper
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Ecosystem
p-ISSN: 2165-8889 e-ISSN: 2165-8919
2012; 2(4): 78-87
doi: 10.5923/j.ije.20120204.06
The Principle of Optimal Biodiversity and Ecosystem Functioning
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-Text HTML
Full-Text HTMLElena N. Bukvareva 1, Gleb M. Aleshchenko 2
1A.N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow, Russian Federation
2Faculty of Geography, Lomonosov Moscow State University, Moscow, Russian Federation
Correspondence to: Elena N. Bukvareva , A.N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow, Russian Federation.
| Email: |  |
Copyright © 2012 Scientific & Academic Publishing. All Rights Reserved.
We propose the principle of optimal diversity of biosystems. According to this principle, the optimal values of inner diversity of biosystems correspond to their maximum viability (minimum extinction probability). We have investigated a mathematical model of a two-level “population-community” system in a fluctuating environment. The subsystems of the lower level are interpreted as populations while those of the upper level are interpreted as a community of one trophic level made up of these populations. The optimality criteria correspond to the maximum effectiveness of resource utilization by the biosystems, which is possible to consider as an index of ecosystem functioning. Оptimal values of diversity depend on the intensity of resource flow and the instability of the environment. optimal species diversity increases in more stable and “rich” environments, while optimal intrapopulation diversity decreases in more stable environments and is independent of the intensity of resource flow. These opposite reactions allow us to make an assumption of the different roles of intrapopulation diversity and species diversity in a fluctuating environment: intrapopulation diversity is the basis of adaptation to environmental instability, while species diversity enables a community to use resources to the maximum and effectively.In general, the results of our modelling agree with empirical biodiversity patterns, giving us grounds to propose the principle of optimal biodiversity as a working hypothesis complementary to other ideas about interrelation between biodiversity and ecological functioning.
Keywords: Optimal Diversity, Intrapopulation Diversity, Species Diversity, Ecosystem Functioning
Article Outline
1. Introduction
- The relationship between biodiversity and ecological functioning has been a focus of ecological research for a long period of time. The results of experimental, observational and theoretical investigations demonstrate that this interrelation is a significant phenomenon and is of crucial importance in nature protection theory and practice[1 - 6]. D. Tilman[1] points out that diversity must now be added to the list of factors that influence ecosystem functioning.In our opinion, extremal principles may lead to considerable benefits in the investigations of interconnections between ecosystem properties and diversity. According to these principles, biosystems have a tendency to reach only such states when their important characteristics associated with the survival, viability and development are extremal (maximum or minimum depending on their positive or negative values), for example, the maximum energy effi ciency of an organism, the minimum mortality in the population, the maximum total biomass of the community,etc.These indicators of viability are called optimality crite ria.Optimized characteristics of biosystems are adjustedsuch as to achieve the extreme values of the optimality criteria.The extremal principles have got wide distribution in biology. There are a lot of examples of their successful application in physiology, biochemistry, embryology, evolution theory, population dynamics, and ecology. However, in the field of biodiversity research, the capacities of this method have not been used in full measure.
2. Principle of Optimal Biodiversity
- In the field of biodiversity researchesthe two following main extremalapproaches are possible.One approach is based on the assumption that the diversity of elements of a biosystem (complexity of a biosystem) is maximized. An example of such approach is the entropy extreme principle for communities[7] which implies the maximization of community complexity at fixed volumes of resource consumption by different species.We suggest the second approach called optimal diversity principle[8]. This principle is based on the suggestion that the diversity of elements of a biosystem is related to the fundamental characteristics which define its viability (survival probability). These vital characteristics have a tendency to reach their maximum given their corresponding value of diversity (Figure 1). This value of diversity is optimal (D* in Figure 1).At each passing moment of time, the system is trying to reach a state with maximum viability and optimal diversity (V*, D*). When the environmental conditions are changed, the system adapts to those and changes its parameters so that the optimal value of its diversity also can be changed. We can assume that the diversity levels of undisturbed natural systems are the closest to the optimal values. An artificial decrease or increase of inner biosystem diversity in line with the fast environmental changes leads to a decrease of biosystem viability.
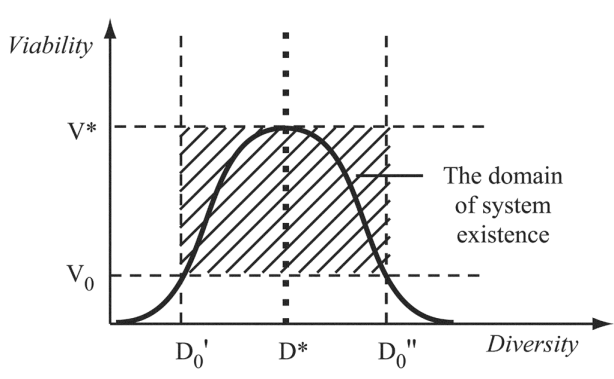 | Figure 1. Optimal value of diversity (D*) corresponds to maximum biosystem viability (V*). V0, a critical value of viability; D0, a critical value of diversity; the shaded area is a domain of system existence |
3. Two-level “Populations-community” Model
- We have demonstrated operability of the principle of optimal diversity by the example of models of two types of biological systems - statistical and structural (in accordance with the notation of the two ways of forming of a top-level control system by A. Lyapunov[9], which can be interpreted as model of phenotypic diversity of the population[10],[11] and the optimal number of species in a community of one trophic level[11].As a next step we have developed and investigated a mathematical model of two-level “populations-community” system in which optimal diversity is forming at both levels during their interaction.Full description and mathematical equations have been presented in previous publications[12]. Here we briefly repeat its basic properties. Еnvironment is characterized by the intensity of resource flow and by the environmental parameter that can be interpreted as any resource characteristic (for example, light wave length, size of the prey and so on) or as any environmental factor that supplies resource consumption (for example, temperature, humidity, etc.). At each passing moment of time, some value of this parameter is realized. The dispersion of the distribution of its values defines the degree of environmental instability.The lower level – population – is represented as the stochastic model which was investigated by means of statistical tests (Monte Carlo method). Populations consist of various phenotypes. The death rate is set by exponential dependence with a constant mortality; reproduction is modeled by a logistic function with birth rate index, which is monotonously decreasing with the growth of population size.Phenotype characteristic is the ability of individuals to propagate in a given environmental conditions (Figure 2). At each passing moment of time, the realized environmental factor f* corresponds with a definite phenotype, for which the given environmental conditions are the most favorable. At this moment, a group of phenotypes breeds around it. The value of dispersion of distribution of breeding at each moment according to phenotypes (black bars in Figure 2) can be interpreted as an index of the width of the zone of individual tolerance. The value of dispersion of distribution of their offspring (shaded bars) serves as an index of diversity reproduced by the population at each step of its development.
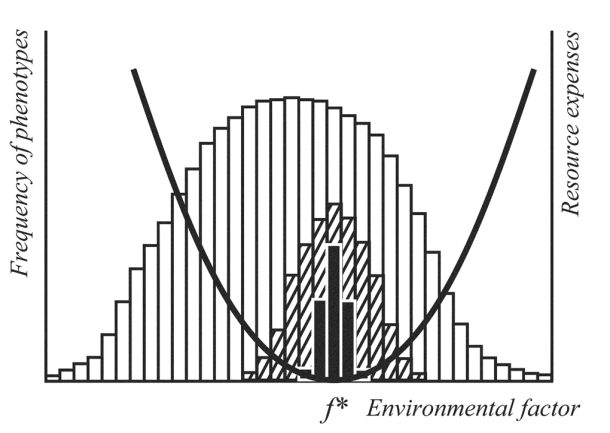 | Figure 2. Phenotypic diversity in population and resource spending by phenotypes. f*, the value of environmental parameter realized at a given moment of time; white bars, existing phenotypes; black bars, phenotypes breeding at a given moment; shaded bars, offspring of the breeding phenotypes, black curve, resource spending by phenotype f* when environment deviates from the f* value |
4. Results of Modeling
4.1. Domain of Population Stability
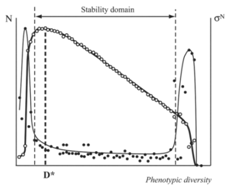 | Figure 3. Dependence of population size N (white circles) and its dispersion σ N (black circles) on phenotypic diversity. Dispersion of population numbers obtained during model tests is an index of population stability (low values correspond to stable populations). D*, optimal phenotypic diversity |
4.2. Existence of Optimal Phenotypic and Species Diversity
- The model experiments reveal the existence of optimal values of phenotype diversity which correspond to the maximum population size/biomass (D* in Figures1 and 3). Any case of diversity deviation from the optimal value leads to a decrease in population size or growth of resource spending. It is interesting to note that the optimal values of diversity in the explored model are close to the bottom border of population stability. If we suppose that natural populations have phenotypic diversity close to optimal values, this result will certainly emphasize the danger of intrapopulation diversity decrease. Even a slight decrease in the level of phenotypic diversity reproduced by the population at each passing moment of time can lead to the loss of its stability. There arise optimal values of species diversity (number of populations in a community) which correspond to the maximum total quantity/biomass of all populations.
4.3. Shift of Values of Optimal Diversity and Population Size Under Changes of Environment
- Optimal values of intrapopulation and species diversity as well as population size depend on the degree of environment stability and the intensity of resource flow in the following way.At the population level:- the optimal values of intrapopulation diversity decrease in more stable environments and are independent of the intensity of resource flow (Figure 4a);- the maximum values of population numbers/biomass increase in more stable and “rich” environments (Figure 4a);- the minimum values of resource spending per individual decrease in more stable environments and are independent of the intensity of resource flow (Figure 4b).At the community level:- the optimal values of species diversity increase in more stable and “rich” environments;
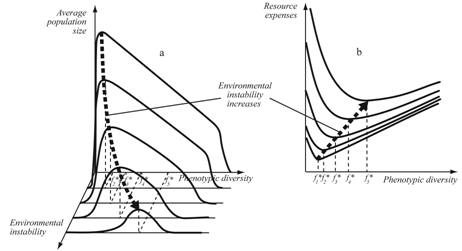 | Figure 4. Optimal values of phenotypic diversity (f*), population numbers and resource spending in environments with different stability |
4.4. Demographic Compensation of Shift of Optimal Diversity Values
- The decrease in mortality, as well as the increase in birth rate and the increase in individual tolerance (diversity of breeding phenotypes at each moment), produces the same effect on location of optimal diversity values as stabilization of environment (Figure 5).
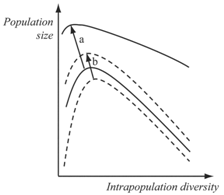 | Figure 5. Changes in optimal values of intrapopulation diversity: a, at increase in fertility; b, at increase in individual’s ecological tolerance |
5. Discussion
5.1. Criteria of Biodiversity Optimality and Ecosystem Functioning
- We used in essence the same optimality criteria at population and community levels: the maximum quantity/biomass at a fixed amount of available resource or the minimum spending of resource at a fixed total quantity/biomass. These criteria are reduced to only one – minimum spending of an individual or biomass unit can be considered an effective measure of resource utilization by the biosystem. The model populations and communities establish the optimal inner diversity at which their effectiveness is maximum. Such an optimality criterion for biosystems seems reasonable enough, because it is directly linked to biosystem viability. The optimality criteria used can give a rough estimate of the effectiveness of ecosystem functioning. Indeed, for stationary communities which use all the resource available, the constantly supported total biomass or effectiveness of resource utilization can be an index for supporting and regulating ecosystem services. These characteristics are often applied as indices of ecosystem functioning in experiments and field observations[13 - 16]. Thus, we may suppose that if a community is in an optimal state, ecosystem functioning is maximum. If a community leaves a zone of optimal diversity values, the effectiveness of ecosystem functioning decreases.
5.2. Possible Mechanisms of Optimization of Diversity
- Possible mechanisms of optimization of diversity throughout ecological, microevolutionary and evolutionary processes are considered by us in a separate publication[17]. Hereweonlybrieflylistthemainmechanisms.Optimization of species diversity in a community is going onin the process of its "self-assembly" from the available regional species pool. The lack of species in the regional pool for any type of extreme habitats may lead to humpbacked function of species number on some environmental gradient (see Section 5.4) or can be compensated by the formation of the intraspecies ecological forms. During succession optimum values change. Climaxcommunity in the framework of our model can be considered as a community that uses every available opportunityto achieve the optimal values of diversity.Optimization of intrapopulation diversity can occur primarily due to changes in the diversity of offspring (shaded bars in Figure 2). This parameter depends on the level of genetic diversity in the population and the average width of the reaction norm. "Tuning" diversity within the reaction norm does not require genetic changes and is the most labile mechanism ofoptimization of phenotypic diversity. When environment stabilizes necessary reduction in intrapopulation diversity can be quickly achieved by producing more homotypic offspring. In moderate destabilization of the environment phenotypic diversity increases due to epigenetic components within the reaction norm.At extreme deviations of environmental conditions offspring phenotypes may go beyond the previous norm of reaction.Further optimization of the phenotypic diversity may also occur due to changes inintrapopulation genetic diversity, but it obviously requires more time.Other population parameters that shape phenotypic diversity - the width of the ecological tolerance of propagatingphenotypes (black bars in Figure 2), the function of the resource expenditures and the maximum rate of population growth - are species traits and their changes occur in the evolutionary time scale.With a lack of species in a regional pool, optimization of diversity can occur through the development of intraspecific sympatric ecological forms. The formation of discrete intraspecific ecological forms fundamentally differs from the increase in diversity of continuous phenotypic distribution. If we consider the ecological structure of a community, in the first case intraspecific forms occupy different niches, in the second casethe single niche expands.Intraspecific sympatric forms can be represented as a dynamic system, constantly tuning parameters of diversity in accordance with changes of environment –when environment stabilizes the number of discrete ecological forms increases, when environment destabilizes this number decreases.Natural biosystems exist in a changing environment. They must continually "tune" their parameters, including diversity, in accordance with the changes taking place. We can assume that natural undisturbed communities and populations existing in historically typical environment are closest to the optimal diversity values. Anysignificant and rapid (exceeding the speed of biosystems adaptation time) environmental changing and disturbance of the biosystems will make them deviate from their optimal state, and their effectiveness and viability will be reduced.
5.3. Opposite Reaction of Optimal Values and Different Role of Intrapopulation and Species Diversity
- The opposite reaction of optimal diversity values on environmental destabilization at population and community levels allows us to make an assumption about their different role in a fluctuating environment: intrapopulation diversity is the basis for adaptation to environmental instability, while species diversity due to niche differentiation enables the community to use resources effectively. Some experiments show that higher species diversity stabilizes and increases the ecosystem processes but destabilizes and decreases the population level[18], and that species diversity increases biomass production but decreases community resistance to drought perturbations[19]. These results indirectly confirm the different role of intrapopulation and species diversity and can be interpreted as a reflection of the fact that the adaptation to environmental fluctuations is carried out primarily at the population level.J. Norberg and coauthors[20] have found out a similar behaviour of model system: in a fast changing environment, phenotype variance increases and total system’s biomass decreases. However, these authors interpret phenotypes as generalized phenotypes of separate species inside a functional group and make up a conclusion about the growth of interspecies differences inside a functional group when the rate of environmental variability increases.The opposite behaviour of optimal values and the probable different role of intrapopulation and species diversity in a fluctuating environment allow us to expand some recent ideas about biodiversity functioning. For instance, a large number of species is considered a kind of community preadaptation and “insurance” against unpredictable environmental shifts[21],[22]. But we hypothesize that the adaptation of communities to a high level of stationary environmental fluctuations increases intrapopulation diversity and decreases species diversity. Our results also allow us to change the angle of view on the question “does functional redundancy exist[23]?” The principle of optimal biodiversity assumes that functional redundancy is the optimized parameter of a community as well as intrapopulation and species diversity as a whole. The degree in which the ecological niches overlap is a result of optimization of diversity parameters at population and community levels. Functional redundancy is not only a “safety factor” similar to engineered redundancy[24], and a reservoir of variations allowing to ad-just to changing conditions[25], but also the optimized property that allows the maximum effectiveness of community in the given environment.
5.4. Does the Optimal Biodiversity Principle Agree to Empirical Data?
- The optimal biodiversity principle predicts that natural communities which are adapted to “rich” and stable environments consist of a large number of species with low intrapopulation diversity (specialists), while communities which are adapted to “poor” unstable environments consist of a small number of species with high intrapopulation diversity (generalists). In “rich” unstable and “poor” stable environments, we expect the medium level of species and, consequently, high and low intrapopulation diversity (Figure 6). We emphasize that these conclusions are made for undisturbed natural systems which exist in a typical historical environment.
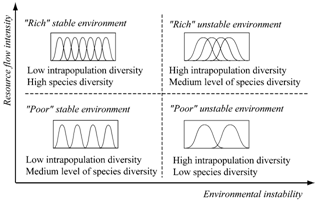 | Figure 6. Assumed levels of species and intrapopulation diversity in communities which are adapted to different environments |
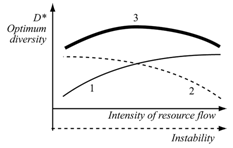 | Figure 7. Changes in optimal diversity values at simultaneous enrichment and destabilization of the environment: 1 - in-crease in diversity at enrichment; 2 - decrease in diversity at destabilization; 3 - the sum curve |
6. “Diversity - Ecosystem Functioning - Environment” Relationship
- Many authors[17],[22],[38] have pointed at bidirectional interrelations between diversity and the main characteristics of ecosystem functioning (stability, magnitude, productivity). Moreover, this is under the influence of environmental conditions – the intensity of available resource flow and the degree of environmental stability (Figure 8a). So we have quite an inoperable scheme where all things are interconnected with each other.The optimal biodiversity principle changes this scheme (Figure 8b) to a two-lewel self-optimizing hierarchical system (populations-community) which adjusts its parameters to the given environmental conditions. Diversity at both hierarchical levels is the optimized parameter, in which optimal values provide maximum resource effectiveness and biosystem viability. Environment parameters (instability and richness) govern optimal diversity values and extreme values of ecosystem functioning indexes. Such a notion may help overcome some obstacles in the practical application of “biodiversity-ecosystem functioning” hypothesis in nature conservation; for example to shift the formulation of biodiversity conservation aims from maximum diversity and maximum ecosystem functioning[39] to optimal ones
7. Biodiversity and the Purpose of Management of Ecosystem Services
- Ecosystem functions may be grouped into three main categories: the formation and maintenance of environmental parameters suitable for human life – environment-forming functions; the biomass taken by humans from nature (seafoods, timber, fodders, fuel, raw materials for pharmaceutics and industry, etc.) – productive functions(so-called ecosystem goods); information present in natural systems and their cultural, scientific, and educational significance – informationfunctions. This division of ecosystem functions differs from that adopted for ecosystem services in the international documents (e.g.[40]), but we propose to use it, because it is more convenient for understanding the biological and other natural processes.The basic characteristics of the biosystems – theirbiomassand levels ofthe internal diversity – used in our model, can act as the indicators of effectiveness. The effectiveness of ecosystemservicesisinextricablyconnectedwith indicators ofbiological diversity, therefore it is necessary to consider the status andpossible changes inbiodiversity for developmentof themethodsand strategies ofthe ecosystem servicesassessment and using. Hence, while determiningthe objectives for management of the ecosystems functions as a single complex,itisnecessarytotakeintoaccountthe changes in biodiversity andbiomass, which will take place if using any given functions (Table 1).
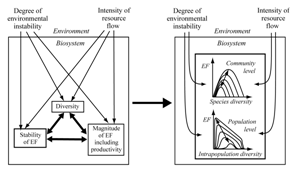 | Figure 8. Relationship between diversity and general characteristics of ecosystem functioning and environment in the context of optimal biodiversity principle |
|
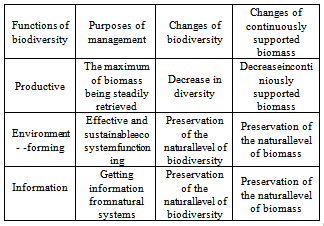
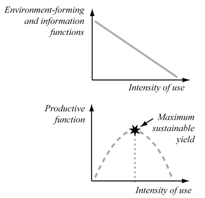 | Figure 9. Alterations of variousbiodiversity functionsunder theintensification of the biological resources exploitation |
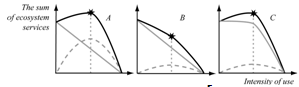 | Figure 10. The value of ecosystem services (S)fordifferent ratiosof productionand theenvironment-formingservices.Graydashed lines-productiveservices; solidgray line-environment-formingservices; solid black line- the total value ofservices;an asterisk-the maximum sustainableyield (explanation in text) |
8. Conclusions
- 1. The proposed principle of optimal biodiversity supposes that the optimal values of inner diversity of the biosystems (intrapopulation diversity and species diversity) correspond to their maximum viability. 2. The results of mathematical modeling have showed the existence of optimal values which obtain maximum effectiveness of resource utilization at the population and community levels. Maximum effectiveness of resource utilization is possible to consider as an index of effectiveness of the ecosystem functioning.3.The optimal values of diversity at the population and community levels depend on environmental instability in an opposite manner: optimal species diversity increases in more stable environments, but optimal intrapopulation diversity decreases. These results speak about the different role of intrapopulation and species diversity: intrapopulation diversity is the basis for adaptation to environmental instability, while species diversity enables a community to use the resource to the maximum and effectively. Thus, the principle of optimal biodiversity integrates population and community levels in the concept of interconnection between biodiversity, ecosystem functioning and environmental conditions.4.The predictions of optimal biodiversity principle agree to general biodiversity patterns and empirical data of experiments and field observations. Seeming contradiction between unimodal (humpbacked) dependence of diversity on productivity and our predictions of its positive form may be explained by species pool hypothesis or by simultaneous enrichment and destabilization of the environment which shift optimal diversity values in the opposite directions. Thus, the optimal biodiversity principle may be proposed as a working hypothesis complementary to other ideas about interrelation between biodiversity and ecological functioning.5.The optimization concept of the “diversity - ecosystem functioning - environment” relationship may be used as a complementary approach in new strategyof nature management.
ACKNOWLEDGEMENTS
- The work has been supported by the “Biodiversity” program of fundamental research of the Russian Academy of Sciences.