-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2021; 11(2): 25-31
doi:10.5923/j.ijaf.20211102.01
Received: Apr. 5, 2021; Accepted: Apr. 28, 2021; Published: May 15, 2021

Seed Viability Trial of Two Balsa Varieties through Direct Sowing in East New Britain
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML1Department of Fisheries, PNG University of Natural Resources and Environment, Kokopo, Papua New Guinea
2Department of Agriculture, PNG University of Natural Resources and Environment, Kokopo, Papua New Guinea
Correspondence to: Kari Iamba, Department of Agriculture, PNG University of Natural Resources and Environment, Kokopo, Papua New Guinea.
| Email: |  |
Copyright © 2021 The Author(s). Published by Scientific & Academic Publishing.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

A viable seed is one which is capable of germination under suitable conditions which also includes dormant but viable seeds. A total of 3600 germinated seedlings were recorded over 22 days (3 weeks). The objective of the study was to determine the potential capacity of the seeds to germinate under double seeding to meet high demand of field planting. Three response variables were tested; double germination, single germination and post single germination. Most doubletons and singletons germinated on day 6. The doubletons were statistically significant on day 6 (347 ± 105, p<0.05). A similar scenario can be seen in singletons on day 6 (668 ± 38.5, p<0.05). Post-singletons appeared in day 8 (53 ± 8.0, p>0.05) with similar germination counts on day 10 (53.5 ± 13.5, p>0.05). The viability of doubletons in Honduras Lancetilla (HL) was higher than those of Vimmy. However, Vimmy did well on singletons and post-singletons giving similar germination counts. From these results, it was recommended that new sowing be done on the second week of the month to ensure a continuous supply of seedling production for the third week (day 8 onwards).
Keywords: Viable, Dormant, Doubleton, Singleton, Post-singleton, Germination
Cite this paper: Boas Malagat, Kari Iamba, Seed Viability Trial of Two Balsa Varieties through Direct Sowing in East New Britain, International Journal of Agriculture and Forestry, Vol. 11 No. 2, 2021, pp. 25-31. doi: 10.5923/j.ijaf.20211102.01.
Article Outline
1. Introduction
- Seed viability can be determined from germinated seeds or prior to planting. A viable seed is one which is capable of germination under suitable conditions which also includes dormant but viable seeds (Koller et al. 1962). It has been shown that, the lower the temperature and moisture content, the longer the period of seed viability (Roberts 1972). For this study, the dormancy was broken through mechanical sclerification before viability was measured by germination (del Tredici & Torrey 1976; Bradbeer 1988; Venier et al. 2012). A non-viable seed would fail to germinate even under all optimal conditions. Ungerminated seeds could be checked for non-viability by tallying any apparently non-viable seeds (i.e., shrivelled caryopsis or lemma/palea without a caryopsis) and by using standard tetrazolium testing procedures (Grabe 1970; Van der Walt & Witkowski 2017). Bradbeer (1988) defined viability has a measure of the suitability of a seed set to produce a satisfactory product. The viability of seeds depends on a number of factors such as dormancy of seed, maturity, quality of seed and environmental conditions where it is sown (Koller et al. 1962; Howcroft 2002). There were two separate trials done to test the viability of two varieties of Ochroma lagopus Swartz: Honduras Lancetilla (HL) and Vimmy. Vimmy is a local variety of East New Britain (ENB) Province while HL originated from Central America. Both varieties were subjected to normal nursery conditions and seed viability was quantified from successful germinations. According to Midgley et al. (2010), the Honduran trees have been proven to perform exceptionally well since an improvement program that began in Papua New Guinea in the 1980s. Seeds of O. lagopus was initially imported into ENB prior to 1938 and subsequently in 1946 by Australian Balsa (Howcroft 2002). Vimmy is believed to be a surviving descendent of the O. lagopus that was initially introduced. The second species, O. pyramidale, was introduced and established at Kerevat in ENB (Whitmore 1969). In 1977, the original silvicultural techniques of balsa in PNG was amended to cater for new requirements in the balsa industry (White 1965). The two species, O. lagopus and O. pyramidale are sometimes used interchangeably (Howcroft 2002). This trial was conducted to improve the nursery system by moving from normal transplanting of seedlings in seed trays to direct seed planting. Transplanted seedlings are prone to injuries to their root systems caused by pressure of surrounding soil particles during the uprooting process. Narrow, shallow holes and mishandling of seedling during the transplanting process may result in ‘J’ roots and also affects normal growth of seedlings resulting in poor development which in extreme cases ends up in loss of seedlings (Londo & Dicke 2005). The objective was to determine the viability of balsa seeds of Honduras Lancetilla and Vimmy varieties through direct sowing.
2. Materials and Methods
2.1. Study Site
- The study was carried out at Takubar nursery site at Kokopo, East New Britain. The experimental site is located approximately at 4°20' 40.91"S and 152°18' 19.52"S at an elevation of 13m. Soil type is sandy loam, well-drained, fertile and calcareous in nature (Iamba & Yaubi 2021). Sandy loam soil in ENB has good binding traits and has proven to be an adequate medium of raising seedlings. With sandy loam soils, there is no need for sterilization except if infection occurs. The climate is classified as tropical with a great deal of rainfall experienced all year round even in the driest month . The climate is classified as tropically humid with an average rainfall of 2780 mm per annum and only mild dry season. The average annual humidity ranges from 77-79% with temperature of 27-29°C (Iamba et al. 2021). These environmental conditions are highly suitable for balsa cultivation (Howcroft 2002).
2.2. Nursery
- Thirty five (36) trays of 50 cones each were filled with sieved pure garden soil media and sown directly with pre – treated balsa seeds. The soil sieve consisted of a square wooden frame of dimension 2.5 x 3.0 m2 with coffee wire fastened at the sides with “u” nails. Seed pre-treatment involved soaking of seeds in boiled water at 80°C for 1-2 hours then soaking in cold water overnight with continued changing of water every two hours until sowing time (Alcântara et al. 2010). Into each cone, 2 soaked seeds were directly planted 20mm apart into approximately 4mm dibbled holes then covered with soil followed by light watering. The seeds were watered 3 times daily starting at 0730hrs, 1100hrs and at 1530hrs. A total of 3600 seeds were sowed into the thirty six trays. Six (6) days later the germinated seedlings were counted and separated into three groups; trays which had double germination, single seed germination and nil germination. Close monitoring and recording of germinated seeds were carried out every 2 days and continued until the seedlings were 3 three weeks old. After every count, the smaller seedlings from the double germination pots including any new germination from the single pots were transplanted to another pot within the same tray. Further monitoring and watering continued on these seedlings until they were three weeks old and ready for hardening out in the open nursery.
2.3. Data Analysis
- All data were analysed using the Generalized Linear Model (GzLM) with Poisson exponential family and log link function in RStudio (version 4.0.3). A two-way ANOVA and linear modelling were not unsuitable for data analysis since the data recorded for the response variables were not normally distributed according to Shapiro-Wilk test (p<0.05). The Akaike’s information criterion (AIC) in Poisson GzLM was lower than the Gaussian family across all tested variables therefore it was chosen as a suitable model for the data analysis (Royer et al. 2020). Graphical analysis of abundance, defoliation and LAI were constructed with ggplot2 package and its dependencies. Separation of treatment (varieties) and days of germination (time) means were done with Tukey Honest Significant Difference test (Tukey HSD). Cross-factor analyses was also done to test the interaction of varieties and time (days) on double germinations, single germinations and post-single germinations. An integration of cowplots and ggplot2 package with facet function produced specific graphs pertaining to the two O. lagopus varieties.
3. Results
3.1. Temporal Germinations of Honduras Lancetilla and Vimmy
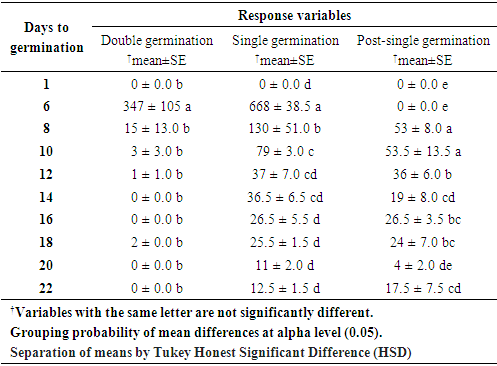
- A total of 3600 germinated seedlings were recorded over 22 days (3 weeks). The objective of the study was to determine the potential capacity of the seeds to germinate under double seeding. The germination counts of the two Balsa varieties varied temporally. Three response variables were tested; double germination, single germination and post single germination. Since the seeds were sown as doubletons, the aim was to assess their germination as a function of time. As shown in table 1, most doubletons and singletons germinated on day 6. The doubletons were statistically significant on day 6 (347 ± 105, p<0.05). However, no significant differences were noted for the other days (p>0.05). A similar scenario can be seen in singletons on day 6 (668 ± 38.5, p<0.05). Day 8 recorded significant germination of singletons (130 ± 51.0, p<0.05) although it was less than that of day 6. Day 10 also had significant germination of singletons (79 ± 3.0, p<0.05) but were lesser than day 8. Day 12 (37 ± 7.0, p>0.05) and 14 (36.5 ± 6.5, p>0.05) had similar singletons with no significant differences. The other days did not record any significant changes in singleton germination. Post-singletons appeared in day 8 (53 ± 8.0, p>0.05) with similar germination counts on day 10 (53.5 ± 13.5, p>0.05). There was a significant drop in singleton count in day 12 (36 ± 6.0, p<0.05). Day 14 (19 ± 8.0, p>0.05) had similar singleton counts as day 22 (17.5 ± 7.5, p>0.05). Day 16 (26.5 ± 3.5, p>0.05) and day 18 singletons (24 ± 7.0, p>0.05) did not differ in germination counts. There was a sharp decline in singletons on day 20 (4 ± 2.0, p<0.05).
3.2. Germination Responses
- Comparing the two varieties, the only statistical difference existed between doubletons of Honduras Lancetilla (HL) and Vimmy. HL recorded higher count of doubletons than Vimmy (46.4 ± 45.07, p<0.05) (fig. 1A). Double germinations of Vimmy was significantly lower than HL (27.2 ± 24.03, p<0.05) (fig. 1A). There was no significant differences between singletons of HL (102.7 ± 67.55, p>0.05) and Vimmy (102.4 ± 60.88, p>0.05) (fig. 1B). Post-singletons also showed no significant difference between HL (24.2 ± 5.53, p>0.05 ) and Vimmy (22.5 ± 7.54, p>0.05) (fig. 1C). The viability of doubletons in HL was higher than those of Vimmy. However, Vimmy did well on singletons and post-singletons.
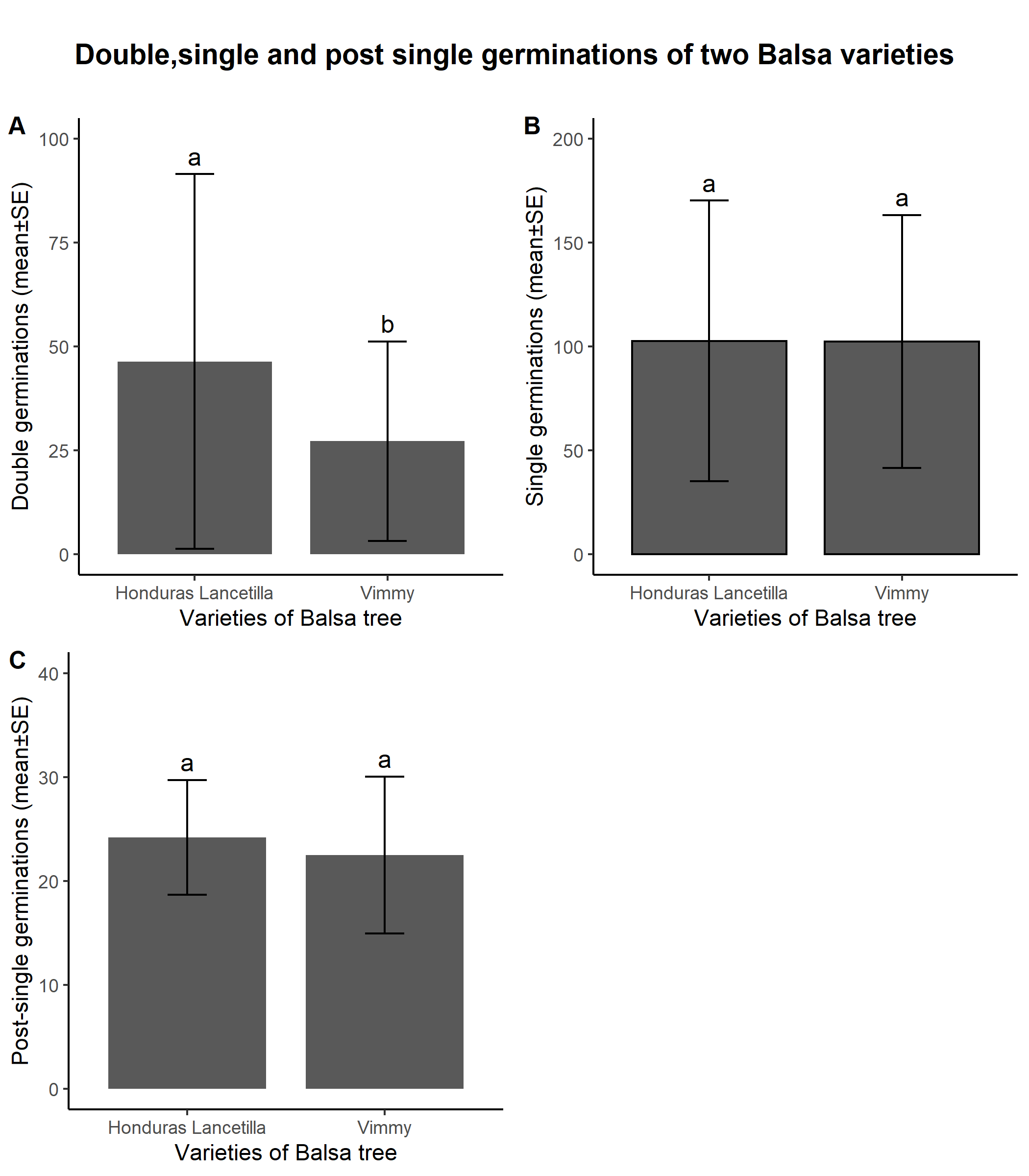 | Figure 1. HL and Vimmy varieties differed significantly under double seeding (A). HL had the highest number of double germinations when compared with Vimmy (p<0.05). Both varieties did not differ under single seeding (p>0.05) (B) and post-single germination (p>0.05) (C). Varieties and variables with same letters are not statistically significant |
3.3. Comparison of Response Variables of Each Variety
- Figure 2 graphically describes the information presented in table 1 on temporal germination patterns. Relative to the two varieties, the germinations peaked at day 6 and then decrease in subsequent days. The total singletons under Honduras Lancetilla (HL) peaked at day 6 then it decreased steeply at day 8 (fig. 2). Subsequent singletons decreased steadily from day 6 to day 22 but was relatively low. Singletons of Vimmy also peaked at day 6 then decreased sharply at day 8 although it was lower than HL. Singletons of Vimmy further decreased at day 10 and day 12 with noticeable drop while subsequent days had rare changes. For doubletons of HL, they peaked at day 6 but dropped steeply in day 8. The subsequent days rarely had double germinations since most seeds have germinated already. The same scenario is also evident from doubletons of Vimmy. Post-singletons of both HL and Vimmy showed similar trend of germination however, Vimmy had slight increase at day 8 and day 10.
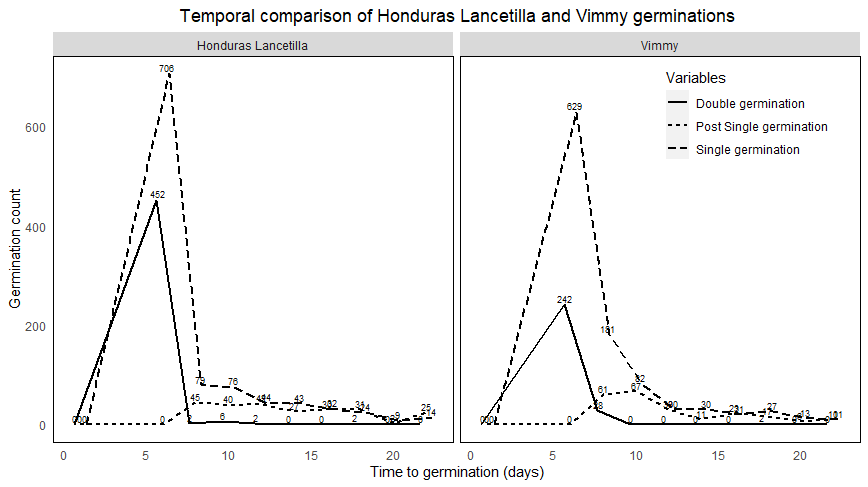 | Figure 2. Single germinations of HL had the highest count at day 6 (p<0.05) followed by double germinations (p<0.05) and post single germinations at very least (p<0.05). Vimmy generally had lower germinations than HL, but followed the same decreasing order as HL. Note that post single germinations are seedlings that germinated singly from double seeding at a later time. Varieties and variables with same letters are not significantly different |
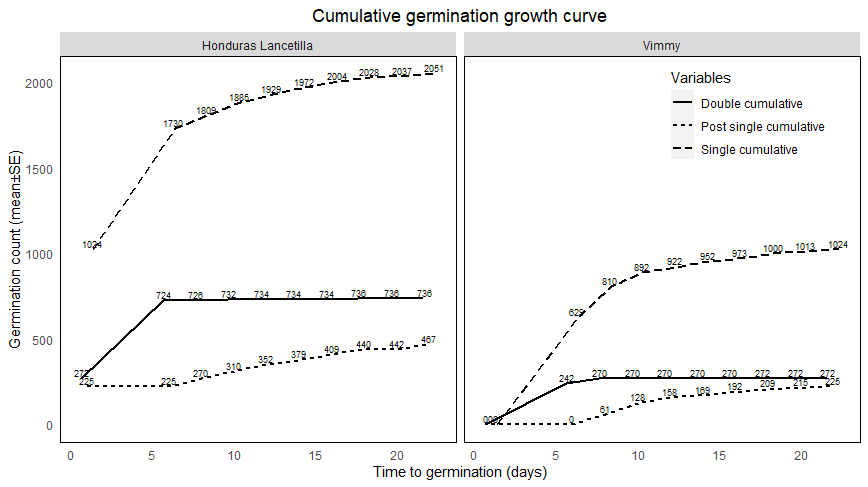 | Figure 3. The cumulative growth curve showed that Honduras Lancetilla (HL) had the highest germination than Vimmy over 22 days (3 weeks). Single germinations from two seeds sown recorded the highest in both varieties followed by double germination and the lowest in post-single germinations (p<0.05) |
3.4. Cumulative Germination Growth
- The cumulative germination curve of singletons and doubletons showed that there was a rapid increase in germinations from day zero to day 6. Subsequent germinations of singletons increased steadily after day 6 while doubletons did not increase at all. For post-singletons, both Vimmy and HL showed slight and steady increase with Vimmy having lower germination counts. Generally, the total germination counts for Honduras Lancetilla were higher than Vimmy variety.
4. Discussion
- High quality seeds of Honduras Lancetilla (HL) was originally sourced from Honduras Seed Garden in Central America where as Vimmy variety was obtained from local seed trees of high profile phenotypic traits in East New Britain (ENB) (Howcroft 2002). Comparing these two varieties, there is a significant difference in doubleton counts (fig. 1A) and at first six (6) days of germination (fig. 2). There were 452 double germinations in HL as compared to 242 in Vimmy. According to Whitmore et al. (2015), Costa Rican variety of O. lagopus has the ability to grow gregariously and as a fast colonizer in secondary clearings of humid neotropics. There are currently limited published information and reports on Vimmy variety although preliminary studies started in 1982 (Howcroft 2002). The difference in doubletons between the two balsa varieties can be attributed to their competitive ability to utilize limited resources such as light, water, temperature, photoperiods and plant regulators (Koller et al. 1962; Metcalfe & Grubb 1997). Differences in the competitive ability of genotypes or cultivars naturally exists in plants (Cousens 1996; Huel & Hucl 1996; O'Donovan et al. 2000). Honduras Lancetilla has higher intraspecific competitive ability meaning even at double seeding, it can still produce two seedlings. In contrast, Vimmy had lower intraspecific competitive ability at double seeding resulting in lower doubletons. As tested in this experiment, altering seeding rate (no. of seeds per hole) is a reliable tool to reduce weed–plant competition than cultivar selection (Korres & Froud‐Williams 2002). Plant varieties that germinate faster and exhibit early vigorous growth have the ability to compete with contender plants like weeds (Bradbeer 1988; Harker et al. 2003). The morphological, physiological and biochemical traits of seeds are also presumed to have contributed to the differences in doubletons, singletons and post-singletons of the two varieties (Lemerle et al. 2001). The rapid and vigorous germination of Honduras Lancetilla could be attributed to its thin coat when compared to the relatively thick coat of Vimmy seeds. Seeds with thick coats are often impermeable therefore breaking seed dormancy can be harder to overcome (Karaguzel et al. 2004). Therefore when HL was subjected to boiling, the seed coat cracked easily allowing imbibition of water that contributes to fast germination. Water plays an important role by activating plant growth enzymes to stimulate root formation to access underground water before growing upwards towards sun and begin photomorphogenesis (Koller et al. 1962). Both varieties were subjected to mechanical sclerification by boiling at 80°C for 1-2 hours to break seed dormancy (Alcântara et al. 2010). As a Central American variety, Honduras Lancetilla have been genetically improved through effective breeding programs (Wycherley & Mitchell 1962; Yu et al. 1982; Alcântara et al. 2010). More breeding work is required for Vimmy to improve its genetic performance. Due to the global significance of ENB, producing 9% of the world’s processed balsa, the balsa industry had to maintain the supply of quality light woods (Jenkin et al. 2019). The aim of the trial was to determine the viability of these varieties and how they could be manipulated to meet the high demand of seedlings required for field planting on a monthly basis. Contracted smallholder growers of balsa accelerated over the past decade to meet the demand and to diversify the income opportunities of farmers since Cocoa, another commodity crop was severely damaged by cocoa pod borer (Conopomorpha cramerella Snellen) (Curry et al. 2015). A modest breeding to improve the genetic quality of ENB balsa germplasm has been realized for some 20 years with irregular program spearheaded by ITTO Balsa Industry Strengthening Project 1996–2003 (Midgley et al. 2010). However, most of these modest breeding programs were done on O. pyramidale while very limited attempt on O. lagopus. Therefore the genetic potential and performance of Vimmy in terms of double germination is much lower than HL. The outcome of this experimental study is a silvicultural approach to boost production. Industrial success of forestry plantation is attained based on effective objectives, which is achieved through appropriate silvicultural management strategies (Leech 2002). Best silvicultural practices depends on site-species selection and choice of seed provenance (Baynes 2005). Seed provenance is a research area that needs attention since most native varieties are well-adapted to the local environmental conditions. These varieties adapts well to two extreme components of the climate, namely the amount and distribution of the rainfall and temperature (Baynes 2005). Therefore, the double germination recorded in the first 6 days is very crucial to meet the monthly field planting target. The double germinations produced by HL variety could play a very significant role in boosting the production of seedlings in the first 6 days of the month. The subsequent single germinations on day 7 for both varieties and also post-germinations of Vimmy towards the end of the second week would ensure a continuous supply of seedlings for extensive field planting.
5. Conclusions
- Direct sowing of balsa seeds (Ochroma lagopus Sw.) into nursery pots resulted in a peak double germination on day six (6) followed by a peak single germination on day eight (8) for both varieties. This shows that the ability of each variety to produce seedlings on the first week presents an opportunity to further manipulate their seeds physiology and growth environment so that more double and single germination can be gained on day 6 and day 8 respectively. The effect of seed coat morphology and physiology could explain the reasons to why HL had a high spike in germination when compared to the local variety. Therefore it is recommended that the varieties should be dipped in boiled water for different lengths of time with Vimmy variety to be dipped for a longer period of time than HL variety. According to the results, it shows that at the end of the third week all germinations were completed. It is therefore recommended that new sowing should be done on the second week of the month to ensure a continuous supply of seedling production for the third week (day 8 onwards). This will maintain a high and steady number of germinations throughout the month. Thereby two direct sowing of seeds per month is recommendable to produce a constant supply of seedlings for field planting.
ACKNOWLEDGEMENTS
- We are grateful for the technical and logistical support of nursery assistants. Special thanks to residents of Takubar for their support throughout the project’s phase.
References
| [1] | Alcântara M, Luz P, Neves L et al. (2010) Evaluation of different methods of dormancy in seeds of breaking Ochroma lagopus (Cav. ex. Lam.) urb. Magistra 22, 141-145. |
| [2] | Baynes JS (2005) The silviculture of small-scale forest plantations in Leyte, the Philippines: a preliminary survey. |
| [3] | Bradbeer J (1988) Seed viability and vigour. In: Seed Dormancy and Germination. Springer. pp. 95-109. |
| [4] | Cousens R (1996) Comparative growth of wheat, barley, and annual ryegrass (Lolium rigidum) in monoculture and mixture. Australian Journal of Agricultural Research 47, 449-464. |
| [5] | Curry GN, Koczberski G, Lummani J et al. (2015) A bridge too far? The influence of socio-cultural values on the adaptation responses of smallholders to a devastating pest outbreak in cocoa. Global Environmental Change 35, 1-11. |
| [6] | del Tredici P, Torrey JG (1976) On the germination of seeds of Comptonia peregrina, the sweet fern. Botanical Gazette 137, 262-268. |
| [7] | Grabe DF (1970) Tetrazolium testing handbook for agricultural seeds. Association of Official Seed Analysts Stillwater. |
| [8] | Harker K, Clayton G, Blackshaw R et al. (2003) Seeding rate, herbicide timing and competitive hybrids contribute to integrated weed management in canola (Brassica napus). Canadian Journal of Plant Science 83, 433-440. |
| [9] | Howcroft N (2002) The balsa manual: techniques for establishment and the management of balsa (Ochroma lagopus) plantations in Papua New Guinea. ITTO East New Britain Balsa Industry Strengthening Project PD 7, 99. |
| [10] | Huel D, Hucl P (1996) Genotypic variation for competitive ability in spring wheat. Plant Breeding 115, 325-329. |
| [11] | Iamba K, Yaubi T (2021) Incorporating lemon grass (Cymbopogon citratus L.) and marigold (Tagetes erecta L.) as non-host barrier plants to reduce impact of flea beetle (Chaetocnema confinis C.) in cabbage (Brassica oleracea var. capitata L.). Acta Entomology and Zoology 2, 95-101. |
| [12] | Iamba K, Yoba S, Wolokom B et al. (2021) Habitat selection by fruit flies (Diptera: Tephritidae) in a tropical agroecosystem in Papua New Guinea. |
| [13] | Jenkin B, Minimulu J, Kanowski P (2019) Improving the smallholder balsa value chain in East New Britain Province, Papua New Guinea. Australian Forestry 82, 23-31. |
| [14] | Karaguzel O, Cakmakci S, Ortacesme V et al. (2004) Influence of seed coat treatments on germination and early seedling growth of Lupinus varius L. Pakistan Journal of Botany 36, 65-74. |
| [15] | Koller D, Mayer A, Poljakoff-Mayber A et al. (1962) Seed germination. Annual Review of Plant Physiology 13, 437-464. |
| [16] | Korres N, Froud‐Williams R (2002) Effects of winter wheat cultivars and seed rate on the biological characteristics of naturally occurring weed flora. Weed research 42, 417-428. |
| [17] | Leech J (2002) Allowing for risk in forward yield planning. Australian Forestry 65, 232-236. |
| [18] | Lemerle D, Gill G, Murphy C et al. (2001) Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Australian Journal of Agricultural Research 52, 527-548. |
| [19] | Londo AJ, Dicke SG (2005) Measuring survival and planting quality in new pine plantations. Southern Regional Extension Forestry, University of Georgia. |
| [20] | Metcalfe D, Grubb P (1997) The responses to shade of seedlings of very small-seeded tree and shrub species from tropical rain forest in Singapore. Functional Ecology 11, 215-221. |
| [21] | Midgley S, Blyth M, Howcroft N et al. (2010) Balsa: biology, production and economics in Papua New Guinea. ACIAR Technical Reports Series. |
| [22] | O'Donovan JT, Harker KN, Clayton GW et al. (2000) Wild Oat (Avena fatua) Interference in Barley (Hordeum vulgare) is Influenced by Barley Variety and Seeding Rate1. Weed Technology 14, 624-629. |
| [23] | Roberts E (1972) Dormancy: a factor affecting seed survival in the soil. In: Viability of seeds. Springer. pp. 321-359. |
| [24] | Royer JE, Tan KH, Mayer DG (2020) Comparative Trap Catches of Male Bactrocera, Dacus, and Zeugodacus Fruit Flies (Diptera: Tephritidae) With Four Floral Phenylbutanoid Lures (Anisyl Acetone, Cue-Lure, Raspberry Ketone, and Zingerone) in Queensland, Australia. Environmental Entomology. |
| [25] | Van der Walt K, Witkowski E (2017) Seed viability, germination and seedling emergence of the critically endangered stem succulent, Adenium swazicum, in South Africa. South African Journal of Botany 109, 237-245. |
| [26] | Venier P, Funes G, García CC (2012) Physical dormancy and histological features of seeds of five Acacia species (Fabaceae) from xerophytic forests in central Argentina. Flora-Morphology, Distribution, Functional Ecology of Plants 207, 39-46. |
| [27] | White K (1965) Silvicultural techniques in Papua New Guinea forest plantations. Department of Forests. |
| [28] | Whitmore JL (1969) Literature review of common tropical trees. Food and Agriculture Organization of the United Nations AGRIS. |
| [29] | Whitmore N, Aplin K, Arihafa A et al. (2015) A rapid biodiversity survey of Papua New Guinea's Manus and Mussau Islands. Wildlife Conservation Society, Papua New Guinea Program. |
| [30] | Wycherley P, Mitchell B (1962) Growth of Balsa trees Ochroma lagopus Sw. at the Rubber Research Institute Experiment Station. Malayan Forester 25, 140-149. |
| [31] | Yu P, Li X, Zou S (1982) A preliminary report on the experiment of introducing and cultivating of balsa tree (Ochroma lagopus Sw.). A preliminary report on the experiment of introducing and cultivating of balsa tree (Ochroma lagopus Sw), 101-107. |