-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2017; 7(5): 103-110
doi:10.5923/j.ijaf.20170705.01

AMMI Analysis of Yield Performances of Argan Genotypes (Argania spinosa L. Skeels) over Disparate Years and Preselection of Promising Trees
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLMohamed Louay Metougui1, Mimoun Mokhtari2, Ouafae Benlhabib1
1Department of Production, Protection & Plant Biotechnology, Hassan II Institute of Agronomy and Veterinary Medicine, Rabat, Morocco
2Department of Horticulture, Horticultural Complex of Agadir, Hassan II Institute of Agronomy and Veterinary Medicine, Ait Melloul, Morocco
Correspondence to: Mohamed Louay Metougui, Department of Production, Protection & Plant Biotechnology, Hassan II Institute of Agronomy and Veterinary Medicine, Rabat, Morocco.
| Email: |  |
Copyright © 2017 Scientific & Academic Publishing. All Rights Reserved.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Argan tree (Argania spinosa L. Skeels) is an emerging oil species of great social and economic interest in Morocco. The thriving national and international demand for its oil increased the anthropomorphic pressure on its forests, slowing the natural regeneration of the natural stands. In order to alleviate the pressure on the species and to satisfy the growing demand for its oil, the domestication of the argan tree and its cultivation should be urgently implemented. High and stable yield over years is an extremely important trait in argan tree selection and should be among the main objectives of its domestication programs. This study aimed to follow-up the production of 122 argan genotypes preserved in situ, at Admine reserve of the Horticultural Complex of Agadir in southwestern Morocco, over three contrasted years. The trees bearing fruits for in any year of the study varied from 50 to 73%. The 52 genotypes with a regular annual production were evaluated for their productivity and yield stability (in number of fruits and almond weight) using AMMI (Additive Main effects and Multiplicative Interaction) analysis. The analysis of variance showed a major genotype-to-year interaction (GYI) effect; the genotype too, had a significant influence, while the year had a negligible effect. Data analysis revealed different responses of each individual between years and subsequently the complexity of this trait. The first multiplicative component of the interaction accounted for 67 and 60% of the sum of squares for fruit density and almond yield respectively. The AMMI1 and AMMI2 biplots, and the AMMI stability value were used to discriminate genotypes with superior and stable fruit yield. The analysis also enabled the identification of potentially productive trees in some specific environmental conditions. Thus, allowing the pre-selection of 12 promising genotypes to constitute the genetic basis of stable and highly productive argan cultivars. Such genotypes need a confirmation of their aptitude for cultivation through further investigation.
Keywords: Argania spinosa, Yield stability, AMMI model, GY interaction, Selection
Cite this paper: Mohamed Louay Metougui, Mimoun Mokhtari, Ouafae Benlhabib, AMMI Analysis of Yield Performances of Argan Genotypes (Argania spinosa L. Skeels) over Disparate Years and Preselection of Promising Trees, International Journal of Agriculture and Forestry, Vol. 7 No. 5, 2017, pp. 103-110. doi: 10.5923/j.ijaf.20170705.01.
Article Outline
1. Introduction
- Argan tree (Argania spinosa L. skeels) is a multipurpose tree that belongs to the Sapotaceae family [1]. This species is endemic to southwestern Morocco, where it occupies an approximate area of 952,000 ha [2, 3]. Its main product is the oil extracted from its almonds, rich in unsaturated fatty acids (80%) and γ-tocopherol, highly prized for its therapeutic, cosmetic, and culinary uses [4]. Argania spinosa plays an important socioeconomic and environmental role in the region. According to the Moroccan independent institution for exports control and coordination, the amount of exported argan oil increased from nearly 100 tons in 2004 to over 1,000 tons in 2015. Despite the importance of the species as an emerging alternative oil species, its natural stands are subject to a constant degradation due to the overexploitation, desertification, and farms expansion over its forests [5]. Given the situation, the domestication and establishment of argan orchards can help to decrease the anthropomorphic pressure on its forest and fulfill the strong demand of its oil.Being a wild forest species, argan trees have a vast underlying genetic diversity, making the genotype environment interaction (GEI) very significant for most traits, which reduces the general association between the genotype and phenotype and make the selection process very difficult [6-9]. In the literature, very few studies have addressed the productivity and selection in argan tree [10, 11], and to our knowledge, none have addressed the yield stability issue.Targeting stable and performant genotypes can be more consistent on a better understanding of the GEI pattern. The analysis of variance (ANOVA) is able to identify the GEI as a source of variation but cannot analyze its intrinsic effects, while the principal component analysis (PCA) cannot present the additive effects for genotype or environment. The Additive Main effects and Multiplicative Interaction (AMMI) model is a hybrid analysis that provides a visual inspection and interpretation of the GEI components [12]. It separates the additive main effects for genotypes and environments using a regular ANOVA analysis and then analyses the interaction effect using the multiplicative model provided by the PCA [12, 13]. The aims of this study are to (i) assess the general productivity of the studied argan genotypes, (ii) assess the yield’s stability of genotypes not presenting an apparent alternate bearing (a year with no production) using the AMMI analysis and to (iii) determine those with both high and stable yields through the contrasted years of the study intended for future argan selection and breeding program.
2. Material and Methods
2.1. Study Site and Plant Material
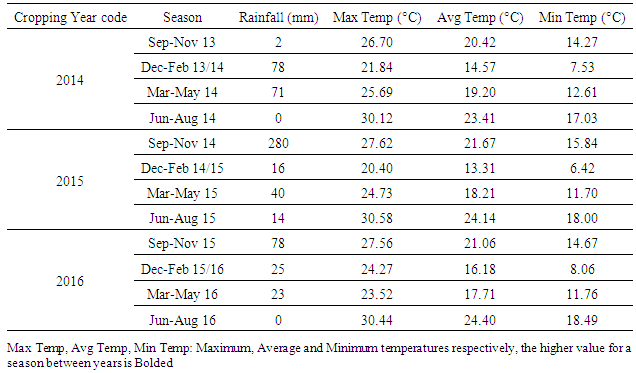
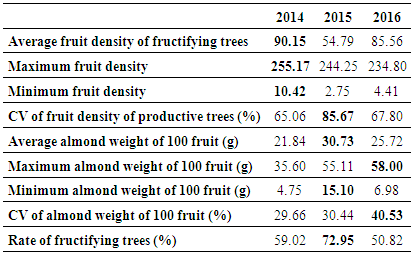
- The study was carried out during three cropping years (from 2013/14 to 2015/16), in the Admine natural reserve placed within the Horticultural Complex of Agadir in upper southwest Morocco. The reserve extends over 28 ha and is located at 9°28’35”W, 30°21’58”N and at an average elevation of 30 m above sea level. The average annual rainfall in the region is 300 mm (2000-2016), the average high temperature varies between 19 and 37°C, and the average low temperature varies from 6 to 21°C. Occasionally, the area experiences winds from the Sahara, which can increase the temperature above 40°C.During the study period, the average seasonal temperatures were comparable between years except for winter 2016, where it was slightly higher than the two previous years (Table 1). Regarding the precipitation levels, 2015 was the wettest year with 314 mm, and with most of the rain (280 mm) occurring during autumn. For 2014 and 2016, both years had slightly more than 100 mm rainfall with the most (78 mm) occurring during winter in 2014 and autumn in 2016 (Table 1).
|
2.2. Yield Estimation
- To compare between trees with different sizes and shapes, the yield was estimated per m² of the productive surface of the tree. For each tree, the total number of fruits present in a 0.25 m² (0.5 m x 0.5 m) of the external tree surface was counted. Then the average density was calculated over 12 values/tree. After fruit ripening, 3 batches of 100 fruits were collected from each tree to estimate the 100 fruits weight and its components, the weight of pulp, shell and almond of 100 fruits. The final yield, used to estimate the productivity and to compare between the different genotypes, was expressed in gram of almonds per external tree surface m².
2.3. Data Analysis
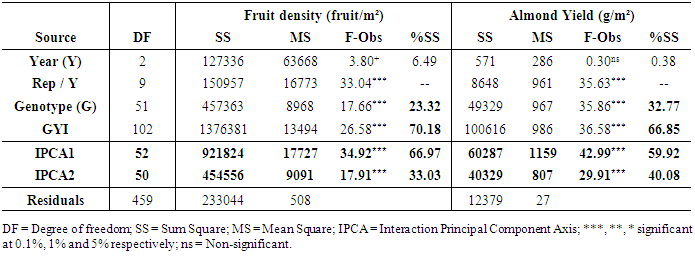
- Rate of productive trees, maximum, minimum, mean and coefficient of variation (CV) were computed for both fruit density and for almond weight of 100 fruit for every year of the study, to visualize the effect of the year over the overall productivity of the sampled trees.Fruit density and almond yield data of the studied genotypes not presenting an apparent alternate bearing tendency (a year without fruit production), were analyzed by the AMMI model to adjust the additive genotype and environmental effects by the analysis of variance. In addition, the adjustment of the multiplicative effects of the G×Y interaction (GYI) was performed by principal component analysis. The AMMI model used in the analyses was as follows [13]:
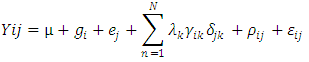 | (1) |
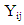 is the fruit density or almond yield of the
is the fruit density or almond yield of the 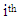 genotype in the
genotype in the 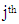 year, μ is the overall mean,
year, μ is the overall mean, 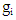 and
and 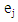 are the fixed genotype effects and environmental deviations, respectively,
are the fixed genotype effects and environmental deviations, respectively, 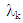 is the eigenvalue of the Principal Component Analysis axis k,
is the eigenvalue of the Principal Component Analysis axis k, 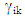 and
and 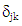 are genotype and environmental factors, respectively, of the singular vectors associated with
are genotype and environmental factors, respectively, of the singular vectors associated with 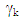 from the interaction matrix,
from the interaction matrix, 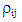 is the residual of GYI, and N is the number of principal components retained.The AMMI Stability Value (ASV) is used to quantify the stability of the genotypes based on the two first Interaction Principal Component Axis and was calculated using the formula developed by Purchase [14]:
is the residual of GYI, and N is the number of principal components retained.The AMMI Stability Value (ASV) is used to quantify the stability of the genotypes based on the two first Interaction Principal Component Axis and was calculated using the formula developed by Purchase [14]: | (2) |
3. Results and Discussion
- The average fruit density was higher in 2014 and 2016 (90.15 and 85.56 fruits/m² respectively), while it was significantly lower (54.79 fruits/m²) in 2015 (Table 2). This decrease in the fruit density was caused by the falling of fruits observed after storm rains, which took place in autumn 2014. Those storms caused a natural thinning, which decreased the fruit density, but helped the improvement of the almond weight. Almond weight reached 30 g/100 fruits in 2015 whereas in 2014 and 2016 it was 21.85 and 25.72 g/100 fruits respectively (Table 2). Such behavior has been reported in other species like avocado, where the excess of water caused premature fruit dropping [15]. In fact, it have been found that the primary effect of flooding is the depletion of soil’s oxygen which usually affects the yield of many species [16, 17].The highest rate of fructifying trees was that of 2015 with 72.95% of the total population, while 2016 presented the lowest rate with only 50.82% of trees producing fruits (Table 2). This high rate in 2015 could be due to the 2014 spring rainfall, which helped the development of new stems and floral buds, and also to the following autumn rainfall that sustained the fruits growth. The difference in the number of productive trees among years could also depend on the alternative bearing ability of the tree genotype, which generally relies more on the endogenous nutritive components or on the flowering-fructification cycle length, in fact it is attested in argan that the period from flowering to fructification stages can vary from 10 to 16 months [18]. Generally, in long cycles, fruit development and flower formation overlap which, probably affect both the development of the old fruits and the ability to form new flow buds. In fact, it has been found in many studies that the presence of developing fruits can inhibit subsequent formation and growth of new ones, because of the difference in sink strength, or the dominance of a one development processes over the other [19, 20].
|
|
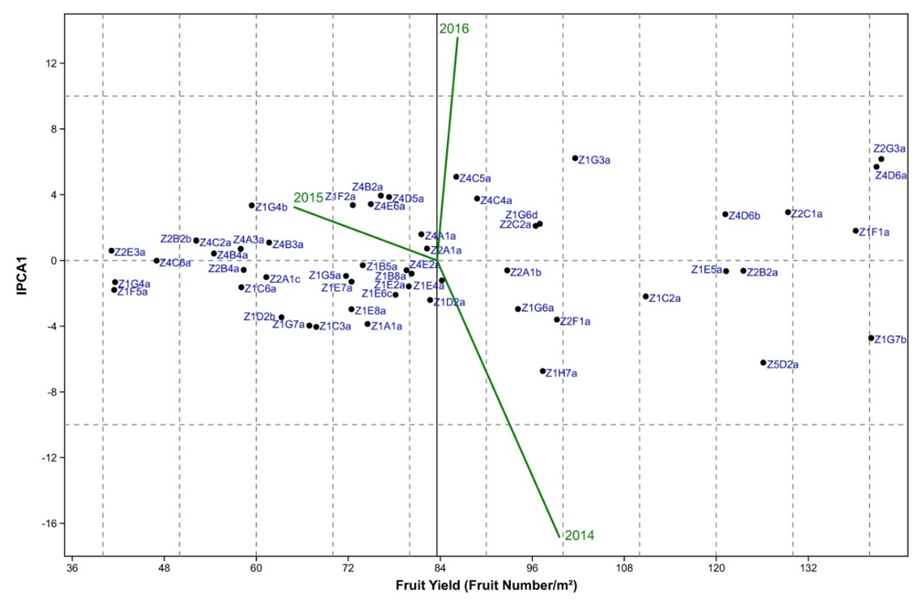 | Figure 1. Interaction principal component axis (IPCA1) against average fruit yield (fruit number/m²) biplot of 52 argan genotypes and three years |
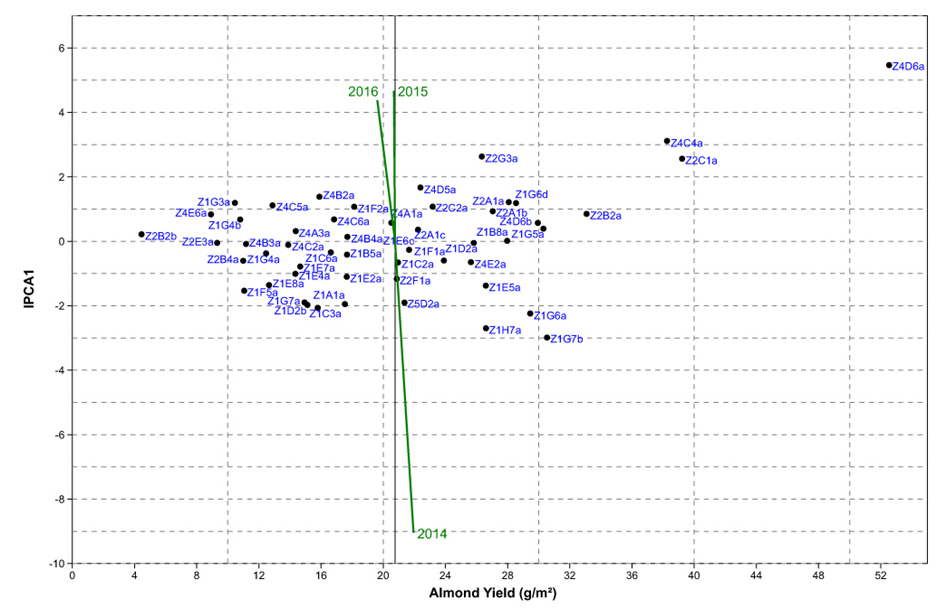 | Figure 2. Interaction principal component axis (IPCA1) against average almond yield (g/m²) biplot of 52 argan genotypes and three years |
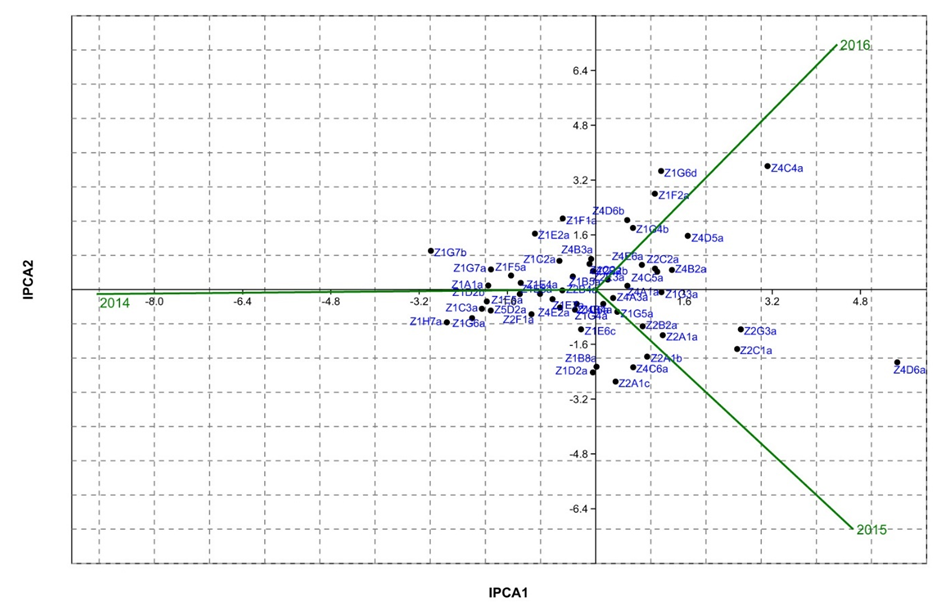 | Figure 3. IPCA1 against IPCA2 biplot for almond yield (g/m²) of the 52 genotypes versus three years |
|
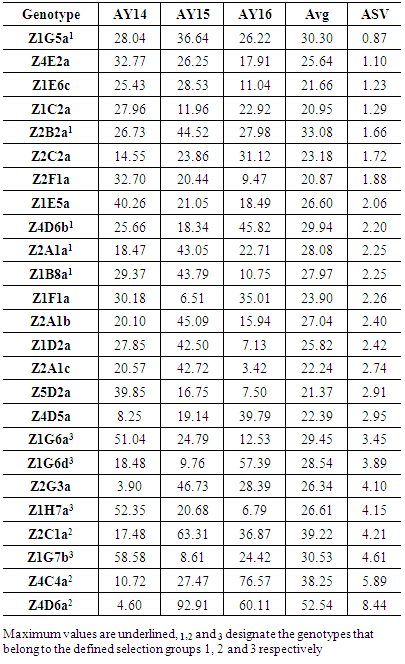
4. Conclusions
- Almond yields data analysis of 122 argan trees in the natural reserve of Admin under natural conditions during the 2014-2016 three-year period showed that the rate of trees bearing fruits varied from 50 to 73%. This investigation study suggests also that argan yield variation is mainly influenced by the genotype-to-year interaction. The study of different yield component, showed that the importance of fruit yield is not always reflected in almond yield, under contrasting environments both traits can be negatively correlated in some genotypes.The AMMI analysis showed that more the genotype is productive, the more its chance to be stable decreases. The use of AMMI model facilitates the selection of stable and high yield genotypes; such genotypes deserve further yield evaluation under orchard conditions.