-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2017; 7(2): 42-51
doi:10.5923/j.ijaf.20170702.02

Morphological Variability, Heritability and Correlation Studies within an Argan Tree Population (Argania spinosa (L.) Skeels) Preserved in situ
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLMohamed Louay Metougui1, Mimoun Mokhtari2, Peter J. Maughan3, Eric N. Jellen3, Ouafae Benlhabib1
1Department of Production, Protection & Plant Biotechnology, Hassan II Institute of Agronomy and Veterinary Medicine, Rabat, Morocco
2Department of Horticulture, Horticultural Complex of Agadir, Hassan II Institute of Agronomy and Veterinary Medicine, Ait Melloul, Morocco
3Department of Plant & Wildlife Sciences, Brigham Young University, Provo, UT, USA
Correspondence to: Mohamed Louay Metougui, Department of Production, Protection & Plant Biotechnology, Hassan II Institute of Agronomy and Veterinary Medicine, Rabat, Morocco.
| Email: |  |
Copyright © 2017 Scientific & Academic Publishing. All Rights Reserved.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Argania spinosa (L.) Skeels is an endemic tree to Morocco, that produce a high-quality oil used for both cosmetic and culinary purposes. In this study, the morphological diversity of argan trees was investigated in an in situ preserved population, at Admine reserve in southwestern Morocco over three years. A total of 122 trees were characterized using 30 quantitative traits. Significant differences among genotypes and among years were seen for all traits. Correlation analysis between traits showed that vigor traits (leaf and shoot sizes) were positively correlated with fruit traits. Broad-sense heritability estimates were high for clustered leaf traits and thorn numbers (H²>0.90) and for most of the fruit, stone and almond traits (H²>0.70). Cluster analysis using Euclidian distances was used to establish the relationships among the argan germplasm and grouped the genotypes into five relatively homogenous clusters. The relationship among the traits was also analyzed by principal component analysis. Highly discriminating traits were those related to fruit, stone and almond morphologies, as well as clustered leaf size, shoot length, and glomeruli density.
Keywords: Argania spinosa, Argan, Diversity, Morphological characterization, Genetic conservation
Cite this paper: Mohamed Louay Metougui, Mimoun Mokhtari, Peter J. Maughan, Eric N. Jellen, Ouafae Benlhabib, Morphological Variability, Heritability and Correlation Studies within an Argan Tree Population (Argania spinosa (L.) Skeels) Preserved in situ, International Journal of Agriculture and Forestry, Vol. 7 No. 2, 2017, pp. 42-51. doi: 10.5923/j.ijaf.20170702.02.
Article Outline
1. Introduction
- Argan tree (Argania spinosa L. skeels) belongs to the monospecific genus Argania and is the only representative of the tropical family Sapotaceae in Morocco [1]. This tree is endemic to Southwest Morocco, where it occupies an area of around 952,200 ha [2]. Outside this major settlement, significant outlying populations are present in the valley of Oued Grou in southeastern Rabat, on the Mediterranean slope of the Beni-Snassen mountains in northern Oujda [3], and in Tindouf region in eastern Algeria [4].Argan is a multi-purpose tree, with leaves and fruit used for livestock feed, and the wood for carpentry and fuel [5]. The primary product of this tree, however, is the highly prized oil derived from its almonds, which has culinary, pharmaceutical, and cosmetic uses [6]. Argan oil contains about 80% unsaturated fatty acids (mainly oleic and linoleic acid) and important levels of γ-tocopherols – a strong antioxidant. Multiple studies have shown the argan oil effect on lowering cholesterol, triacylglycerol, and cardiovascular risks [7, 8]. It has also been shown that argan oil has hypotensive and antiatherogenic properties [7-9].Argan oil is one of the most expensive edible oils in the world, reaching a retail price of 300$/L in the US and European markets where it’s the subject of several cosmetic patents [6]. According to the Moroccan independent institution for export control and coordination, the export of 1000 tons of argan oil in 2015 was worth nearly 20 million US dollars. For the local population in the arid zones where argan grows, the tree plays an important socio-economic role. Indeed, it is estimated that nearly 19% of the local population’s income depends on this tree, with major processing activities, including oil extraction, being traditionally accomplished by women. Thus the argan tree represents an invaluable resource which merits intensive study and preservation [10].Several studies described the genetic diversity seen within argan tree. The first studies dating from the 1990s, showed a strong geographical structure of argan populations using isozymes and chloroplast DNA polymorphisms [11, 12]. Based on fruit and stone morphological variability, Bani-Aameur and Ferradous [13] reported the existence of large variances within populations in contrast with low inter-population diversity. Ait Aabd et al. confirmed the high within population diversity using fruit traits and oil content and reported for the first time the heritability of those characters [14]. These studies were limited to fruit related traits and didn’t consider the existing variability between genotypes in the vegetative part (leaf and shoot traits).Understanding the true genetic diversity within a species is critical for developing in situ and ex situ conservation strategies, which are desperately needed to protect the variability and fecundity of this species. Such studies also provide the basis necessary for developing core breeding germplasm collections used to initiate plant improvement programs. Here we present a study of genetic diversity within the Admine reserve in upper Southwestern Morocco based on the morphological characterization of over 100 argan genotypes using 30 quantitative fruit and vegetative characteristics. The aim of the present study was to determine the amount of genetic diversity within this in situ preserved population, quantify the variability levels of each trait, and identify the most significant discriminating traits in the species.
2. Material and Methods
2.1. Study Area
- The study was conducted at the Admine Natural Reserve of the Horticultural Complex in Agadir, in upper southwestern Morocco. The reserve extends over 28 ha and is located at 9°28’35”W, 30°21’58”N (Figure 1) at an average elevation of 30 m above sea level. The study area is a natural argan forest which has been protected from human pressure for more than 30 years. The area features the subtropical-semiarid climate with warm summers and mild winters which are optimal for argan growth. The average annual rainfall in this region is 300 mm (2000-2016). During the study period, rainfall varied from a minimum of 10 mm (2013) to a maximum of 431 mm (2014).
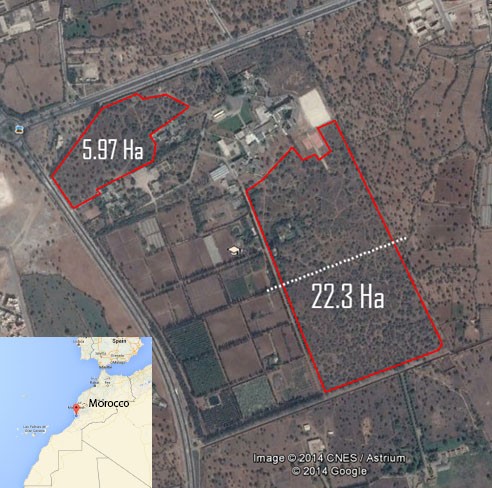 | Figure 1. Map showing the study area |
2.2. Plant Material
- The study was carried out during three years from 2013 to 2015. The Admine Natural Reserve area was divided into 2500 m² plots, from which one tree was randomly selected. A total of 122 trees were studied, covering all the area, and representing nearly 10% of the total number of trees of the reserve. All the studied trees were mature (over 20 years old) and vigorous, with no apparent disease symptoms.
2.3. Morphological Descriptors and Statistical Analysis
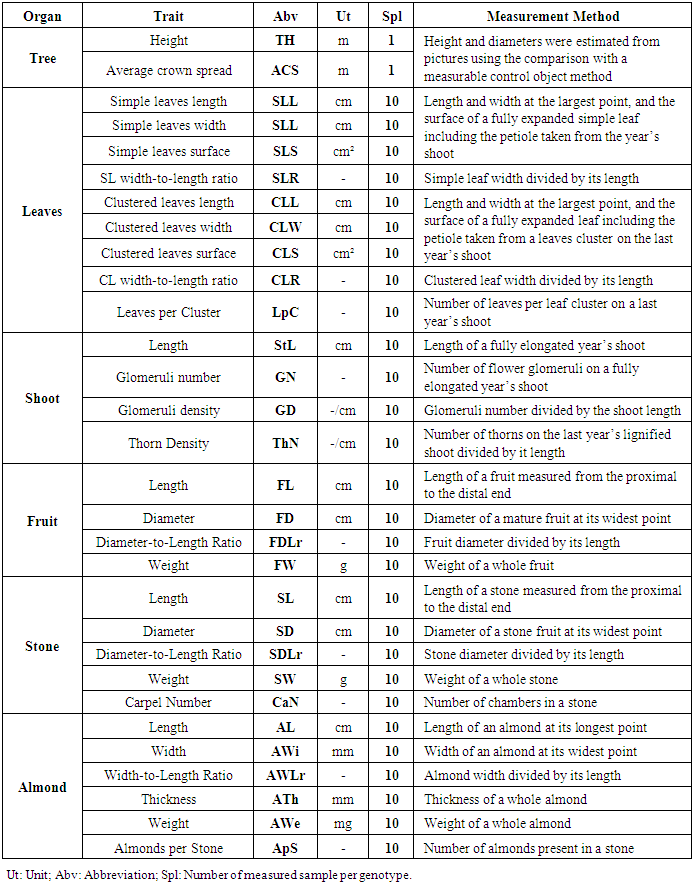
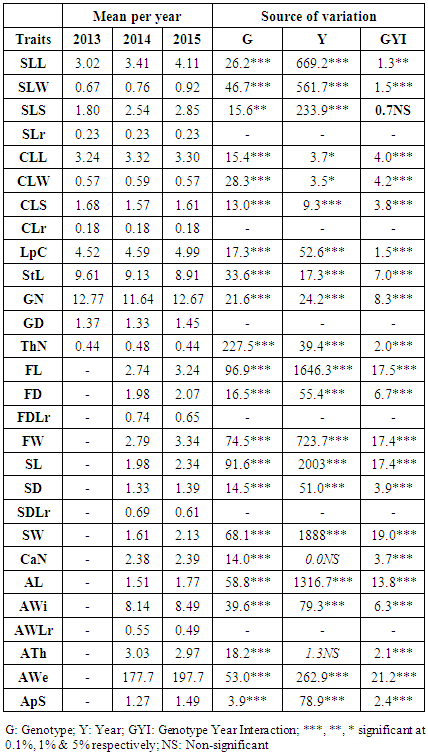
- To assess diversity within the studied population, we used a total number of 30 quantitatively scored morphological traits (Table 1). The characterization included tree, leaf, shoot, fruit, stone, and almond traits. Leaf and shoot traits were recorded for all three years of the study; fruit, stone and almond traits were recorded in 2014 and 2015; while tree height and diameter were recorded only once in 2013.
|
2.4. Genetic Estimates
- Genotypic (σ2g) and Phenotypic variances (σ2p) were calculated using the mean square from the ANOVA tables as formulated by Comstock and Robinson [16]:
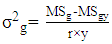 | (1) |
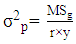 | (2) |
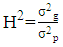 | (3) |
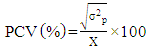 | (4) |
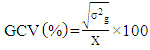 | (5) |
3. Results
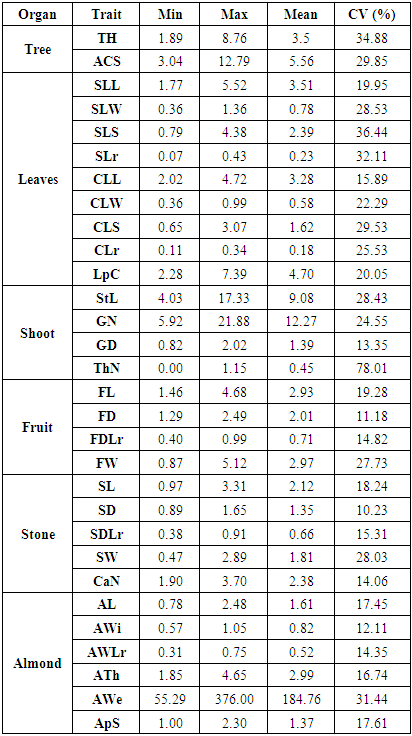
- Mean, minimum, maximum and coefficient of variation (CV) values for the studied characters are given in Table 2. The highest CV identified was for thorn density (78.01%) due to the skewness of its distribution. The next highest CV was that of the simple leaf surface (36.44%), followed by tree height, simple leaf width/length ratio, and almond weight. The lowest CV was that of the stone’s diameter (10.23%). Tree height and crown spread varied widely with CVs of 34.88% and 29.85%, respectively.
|
3.1. Leaf Traits
- Argan trees usually present two types of leaves: simple leaves that grow on newly formed shoots and clustered leaves that grow on older woody shoots. Simple leaf area ranged between 0.79 to 4.38 cm², with an average length of 3.51 cm, width of 0.78 cm, and a surface of 2.4 cm². Clustered leaves were grouped in 2 to 9 leaves per cluster with a leaf area ranging from 0.65 cm² to 3.07 cm² and an average area of 1.62 cm². The CVs of simple leaf dimensions were consistently higher than those of clustered leaves (Table 2). Clustered leaf width/length ratios were equal to 0.18, which were lower than the simple leaf ratio (0.25).Leaf size varied over years. Specifically, there was a better growth of simple leaves in 2015 than in 2013 as illustrated for leaf length (4.11 vs. 3.02 cm), leaf width (0.96 vs. 0.67 cm) and leaf area (2.85 vs. 1.8 cm²) (Table 3). The clustered leaf size variation was lower, varying respectively in its length, width and area from 3.24 to 3.32 cm, 0.57 to 0.59 cm and 1.57 to 1.68 cm² over the three years.
3.2. Shoot Characterization
- The length of new shoots varied widely between genotypes, ranging from 4.03 to 17.33 cm. Glomeruli number ranged between 5.92 and 21.88 per shoot, and from 0.82 to 2.02 per cm (Table 2). Over the years evaluated, 2013 presented the best growth in term of stem length and number of glomeruli/stem, with 9.61 cm and 12.77 glomeruli, respectively, while 2015 had the highest glomeruli density (1.45 per cm).Thorn density on woody shoots ranged between 0 (spineless trees) to 1.15 thorns/cm. The density was slightly higher in 2014 (0.48 thorn/cm) than 2013 and 2015 (0.44 thorn/cm; Table 3).
|
3.3. Fruit Characterization
- Fruit, stone, and almond sizes were measured during two seasons, 2014 and 2015. The fruit weight varied extensively from 0.8 to 5.12 g. Fruit size varied among genotypes from 1.46 to 4.68 cm in length and 1.29 to 2.49 cm in diameter. Stone weight ranged between 0.47 to 2.89 g, and almond weight ranged between 55.29 to 376.00 mg. Stone and almond size showed comparable growth as the fruit and ranged for stones from 0.97 to 3.31 cm in length and 0.89 to 1.65 cm in diameter. The highest almond size was 2.48 cm in length, 1.05 cm in width and 4.65 mm in thickness. The smallest almond size was 0.78 cm, 0.57 cm, and 1.85 mm in length, width, and thickness respectively (Table 2).The fruit diameter-to-length ratio averaged 0.71 and varied between 0.4 and 0.99, respectively for round and apiculate/fusiform type fruits. Thus, this argan population is made up of 79% oval and drop shaped fruits (ratio of 0.60-0.85), 7% round fruits, and 14% apiculate/fusiform fruits.Stone diameter and length were positively correlated (r=0.86), with the diameter-to-length ratio following a similar distribution as the fruits diameter-to-length ratio. Almond width-to-length ratio average 0.52, ranging from 0.31 to 0.75 and showed a positive linear correlation with the fruit ratio (r=0.72). CVs were similar across all fruit characteristics, with the highest CV value for the fruit weight and the lowest for the diameter-to-length ratio. Fruit, stone, and almond size, as well as stone weight were higher in 2015 than 2014. Almond thickness did not show a significant difference between the years (Table 3). Lastly, the number of chambers per fruit stone ranged from one to five and contained one to four almonds, with an average of 2.38 chambers and 1.37 almonds per stone.
3.4. Analysis of Variance and Correlation
- The ANOVA showed highly significant (p<0.01) differences between genotypes (G) for the 22 tested morphological traits, as well as statistically relevant differences between years (Y) except for the number of chambers and almond thickness. The GY interaction was significant for most of the traits except for the simple leaf surface (Table 3).Variance components, which represent the factors’ individual contribution to the total variation, revealed that the genotype was the dominant factor. Its contribution to the total variation ranged between 14.18% for the number of almonds/seed and 89.79% for thorns density (Figure 2). The number of almonds per seed differed slightly between trees, but with considerable variation among trees (the residual represents 71.87% of ApS total variance).
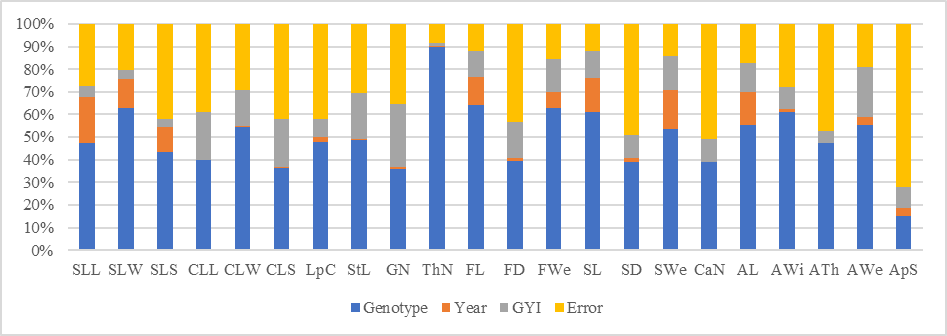 | Figure 2. Contribution of sources to total variance |
3.5. Genetic Estimates
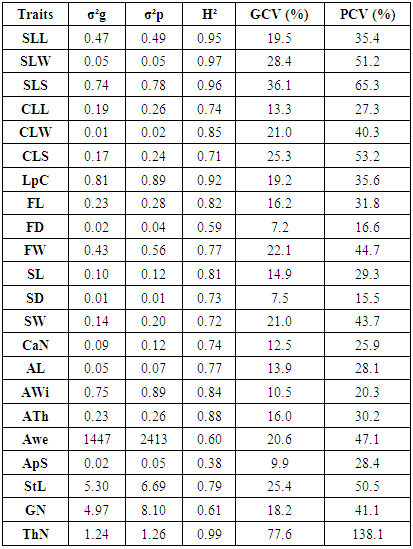
- Genotypic and phenotypic variances, broad-sense heritability, genotypic and phenotypic coefficients of variation of 22 traits are reported in Table 4. The results revealed substantial phenotypic and genotypic variances among trees for all the traits. For most characters, a large proportion of the phenotypic variation was accounted for by the genetic component, highlighting the importance of genetics on the expression of these characters. Heritability was greater than 0.59 for all traits except for ApS, which showed a heritability value of 0.38.
|
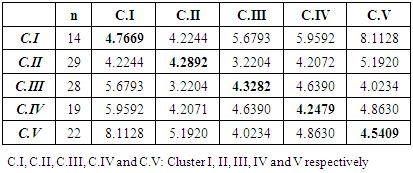
3.6. Cluster Analysis
- The genetic variability among genotypes was studied to know the extent of divergence between the genotypes and gather them into relatively homogeneous clusters. The dendrogram summarizes the phylogenic relationship among 112 genotypes (Figure 3).
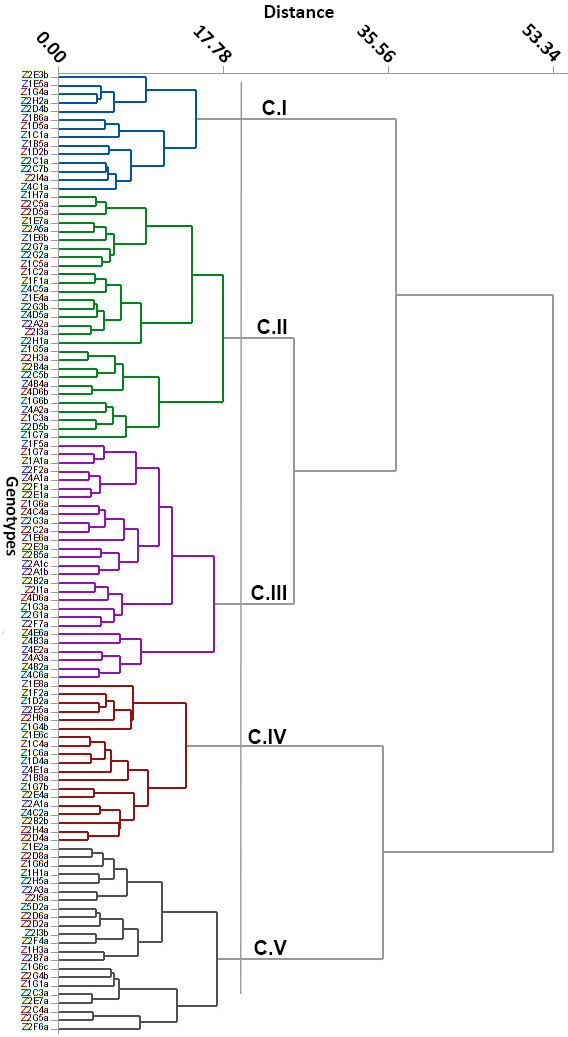 | Figure 3. Genotypes’ clustering using ward linkage and Euclidean distance |
|
|
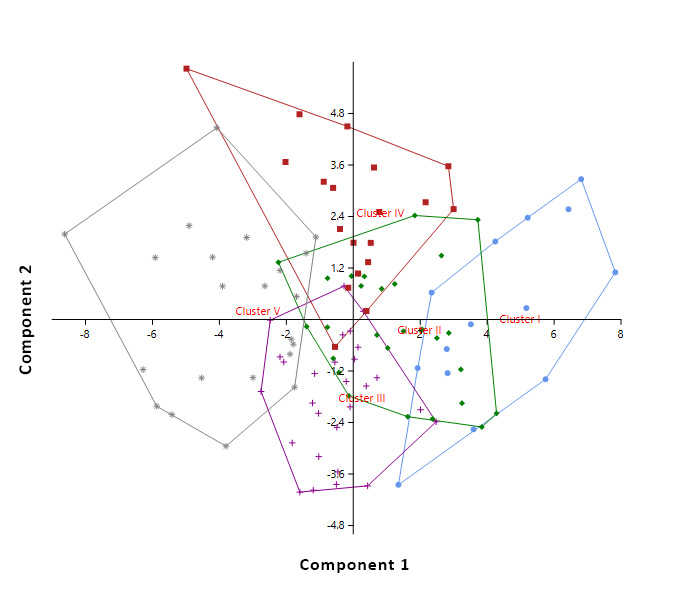 | Figure 4. Two-dimensional PCA plot based on the 2 first components of the studied genotypes with their corresponding cluster (clusters are distinguished with the same colors as in the dendrogram) |
4. Discussion
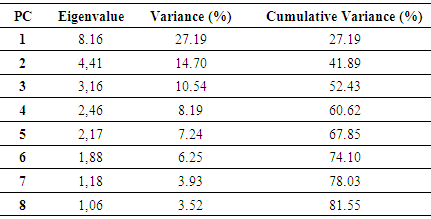
- The understanding of the genetic structure of populations and the study of genetic parameters is essential for effective management of genetic resources and is the first step toward any domestication process. Here we study an in situ protected Argan population growing in the Admine natural preserve of the Horticultural Complex of Agadir. The collected data suggests that a significant phenotypic variability exists within the collection. This high within-population variability is promoted in argan because of its allogamy and entomophily [19, 20]. Variability within core germplasm collections is essential for any domestication work. Our data showed significant differences (p<0.05 to p<0.001) between genotypes for all traits, and between the years for most traits. These results are in agreement with those found within other argan populations [13, 14, 21]. Leaf size values were higher than those reported by Zahidi and Bani-Aameur for both simple and clustered leaves, except for width-to-length ratios which showed the same values [22]. A higher variability was also recorded within our population for simple and clustered leaf width and width-to-length ratio. Fruit, stone and almond weights and dimensions were similar to those reported in previous studies [13, 21, 23]; but our CVs were higher than those reported by Bani-Aameur and Ferradous [13]. The collected data illustrate the high diversity of the Admine reserve population. Such considerable diversity may be related to the protected status of the study area, which helps the trees to develop without any human or animal pressure.Significant variation between years was seen for most traits and could be related to the climatic difference between years, particularly rainfall. In fact, leaf, fruit, stone, and almond traits were greater in the cropping year 2015 when rainfall reached 350 mm, which was higher than the previous years (246 and 151 mm respectively for 2013 and 2014). Similar conclusions were made by Bani-Aameur and Ferradous, where they found an important year effect over fruit traits [24]. Interesting, the climatic difference did not seem to affect the qualitative traits (i.e., general shapes of fruit, stone, and leaves).The correlation matrix between characters indicated that vigor related traits, including shoot length, simple leaf area, number of glomeruli, fruits, kernel and almond sizes were positively correlated. Such correlations between the vegetative organ of the tree and the size of the fruits were reported for several woody species and underscore the role photosynthetic organ play in increasing fruit size and weight [25-27]. The year’s effect on the morphological traits allowed the estimation of heritability. Most of the traits had a broad-sense heritability coefficient greater than 0.60. The magnitude of this coefficient indicates the reliability with which a genotype can be evaluated through its phenotype. Globally, leaf traits had higher heritability than fruits related traits. This is mainly due to the time needed to develop fully expanded leaves and mature fruits [28, 29]. The last one being longer is consequently more affected by the climate.The heritability was maximal (H²> 0.90) for simple leaf characteristics, leaf number per cluster, and spine density which indicate the stability of those characters over disparate years. For, fruit, kernel, almond and clustered leaf traits, they had moderate to high heritability (H² between 0.59-0.89); this suggests that even with a significant environmental effect these traits should respond positively to selection. The number of almonds per stone presents a low H² value and thus is likely strongly influenced by the environment, especially the relative humidity and its effect on the pollination success. The relatively low heritability level of this trait in comparison with the rest of fruit traits is in agreement with previous studies [21, 30].The genetic divergence, based on Euclidean distances, has split the argan collection into five homogeneous clusters; two of the five, Cluster I and IV, present a promising agronomic potential with essentially a high kernel weight. In the principal components analysis, the two first principal components explained only 41.89% of the total variance but they were able to differentiate four clusters over the five ones established through the genetic divergence study; that is because of the relatively high contribution of few traits in the genotype differentiation. The two first PC also showed that fruit and kernel traits are highly discriminating, while leaf traits were of less importance in classifying genotypes.
5. Conclusions
- The main outcome of this agro-morphological characterization is the high variability that exists within the argan collection of Admine reserve. The broad phenotypic diversity detected within this gene pool should be used in selection/breeding programs and to establish a core collection. In fact, the argan conservation, selection, and commercial production are necessary to reduce human pressure on the forest and to prevent the genetic erosion of the species.The morphological characterization revealed significant correlations between the trees’ vegetative traits and the fruits’ size; this will facilitate the early selection of productive genotypes. Most studied traits had a high heritability values and could be efficiently used for argan descriptors establishment. The study also showed the relevance of the fruit traits in differentiating between genotypes. This investigation should be complemented by molecular characterization to fully elucidate the genetic structure of the species. The molecular tools in argan can be also used in marker-assisted selection to accelerate the emerging breeding programs.