-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2015; 5(1): 10-16
doi:10.5923/j.ijaf.20150501.02
Effects of Soil Substrate and Nitrogen Fertilizer on Growth Rate of Acacia senegal and Acacia sieberiana in North Eastern Uganda
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML1Natural Resource Management Programme, Nabuin Zonal Agricultural Research and Development Institute, Soroti, Uganda
2Department of Crop Production Ecology, Swedish University of Agricultural Sciences, Uppsala, Sweden
Correspondence to: Moses Otuba , Natural Resource Management Programme, Nabuin Zonal Agricultural Research and Development Institute, Soroti, Uganda.
| Email: |  |
Copyright © 2015 Scientific & Academic Publishing. All Rights Reserved.
Low tree production as a result of soil nutrient depletion is a major challenge in Uganda. Trees are often planted in marginalized land characterized with very low soil nutrients or absolutely no soil nutrients. The farmers practice poor farming methods such as continuous cultivation and monoculture. The farmers especially smallholders rarely use inorganic fertilizers due to their associated high cost involved. An experiment was set up in North Eastern Uganda to investigate the effects of soil substrate and nitrogen fertilizer on the growth rate of Acacia senegal and A. sieberiana. It aimed at determining the effects of soil substrate and nitrogen fertilizers on the growth rate of A. senegal and A. sieberiana seedlings. A multi-factorial experiment design was used for collecting the data to address the objective of this study. Soil N was determined by Kjeldahl method, P by spectrophotometric method and K by Flame photometry method. The seedlings were subjected to four rates of N treatments (0, 50, 100, 150 mg per plant) as a single dose of ammonium nitrate at the start of experiment and using a randomized complete block design. Two-way analysis of variance (ANOVA) was used to test if the magnitude of effect of soil substrates on growth rate and leaf nitrogen concentration varies between the two acacia species. Pearson correlation analysis was used to test if the leaf N concentration functionally relates to the growth. There was a significant effect (P ≤0.05) of the soil substrate and the species- soil interaction on the growth rate between two acacia species. The mean relative leaf length of A. sieberiana (0.013 mm mm-1 d-1) in unfertilized (N0) soil A was higher compared to those in the soils treated with N fertilizer. While there also was no significant effect (P ≤0. 05) of species, soil substrate and species- soil interaction on the relative stem, leaf biomass growth and the relative root biomass growth at final harvest, there was a significant species effect (P≤0. 05) on the leaf N concentration. There was no statistically significant effect (P≤0. 05) of the leaf N concentration on the growth traits of the two acacia species in all treatments. It can be concluded that unfertilized soil substrates increased the growth rate of A. senegal and A. sieberiana seedlings more than N fertilizer treatments.
Keywords: Trees, Production, Ammonium nitrate, Nutrients, Seedlings, Semi-arid areas
Cite this paper: Moses Otuba , Martin Weih , Effects of Soil Substrate and Nitrogen Fertilizer on Growth Rate of Acacia senegal and Acacia sieberiana in North Eastern Uganda, International Journal of Agriculture and Forestry, Vol. 5 No. 1, 2015, pp. 10-16. doi: 10.5923/j.ijaf.20150501.02.
Article Outline
1. Introduction
- Acacia tree resources are widely distributed in the arid and semi-arid parts of tropical Africa, from Senegal and Mauritania in the west to Eritrea and Ethiopia in the North-East and to South Africa [22]. The acacia species grow well on the dry and rocky hills, and in low-lying dry savannas with annual rainfall of approximately 250 - 350 mm. They also tolerate a maximum temperature of 50℃ and a minimum temperature close to 0℃ and soil pH of approximately between 5.0 and 8.0 [5]. Despite the unfavorable growth conditions for most plants in the arid and semi-arid areas, these acacia species are appreciated in these areas because of their specific morphological and physiological attributes enabling them to cope with those conditions. These trees are used by the rural communities and manufacturing industries in many ways, for example A. senegal to restore soil fertility in rain-fed sorghum-producing areas consisting of clay soils in the Blue Nile region, Sudan [19]. According to [19] also state that these trees can be used for rotational bush – fallow systems. The rotational system consists of relatively short periods of crop cultivation followed by longer periods of fallow under mainly naturally regenerated A. senegal trees when soil fertility declines.The gum exudates tapped from A. senegal and A.sieberiana also contribute to environmental rehabilitation and desertification control through stabilization, reduction of surface run-offs and sheet erosion and soil micro-climate improvement in the Kordofan region of Sudan [6]. In addtion the trees are used as fencing materials, fuel wood, poles, fibre, crafts, medicine and tannins in Uganda.Given the major role of A. senegal and A. sieberiana in the development of local communities in Uganda specifically and globally in general, there are plans to invest into these two species with the aim to increase growth rate and the production of gum exudates [7]. Increased use of the two species could be an opportunity for alleviating poverty in the rural areas particularly in the semi-arid areas.However, recent studies show that the productivity and fertility of soils in the world are declining due to degradation and intensive use of soils without a consideration of proper soil-management practices [11, 5]. It is estimated that around 60% of cultivated soils limit plant production [5].Several factors such as soil type, farmers’ practice, crop residues and mineral fertilizer management influence plant growth and yield [3]. Among those factors, soil texture and the chemical composition of soils remain the major constraints to crop production in large scale in the tropical regions of Africa [18]. This shows that in the coming decades, plant-nutrition - related research will be a high- priority research area contributing to crop production and sustaining soil fertility. Nitrogen, P, and K are the most limiting nut rients to plant growth and crop yield [17]. Nitrogen is primarily required for increasing plant growth and crop yield more than any other nutrients. The absence of N in the plants is often associated with slow growth, reduced leaf size, yellowing; short branches, premature fall colour and leaf drop, and increase the likelihood of some diseases. On the other hand, high amounts of these nutrients often result in excessive shoot and foliage growth, reduced root growth, reduced fruit quality, low plant food reserves, and increased susceptibility to environmental stresses and some plant diseases [8]. Excess N can also lead to an accumulation of nitrate in the edible foliage of plants such as spinach and forage.Although the productivity of most trees are well documented, studies on effects of soil substrate and nitrogen fertilizer on growth rate of A.senegal and A. sieberiana under the operational nursery conditions in Uganda are scanty. To bridge these gaps in knowledge, it was necessary to conduct this study so that more knowledge is generated on the nutrient requirements of these two species. An assessment of soil substrate and nitrogen fertilizer therefore will contribute to equip the stakeholders with the relevant information for improved silvicultural management practices under the nursery operations. It was also hypothesized that (i) the magnitude of effect of soil substrates on the growth rate and leaf nitrogen concentration varies between the two acacia species and (ii) the leaf N concentration is functionally related to the growth traits of the two acacia species.
2. Materials and Methods
- Seeds of A. senegal and A. sieberiana and soil samples were collected from Moroto and Kotido districts in North Eastern Uganda, East Africa (Figure 1). The region is located between latitude 1° 30’– 4° N, longitude 33° 30’– 35° E at an altitude; of 1400 m above sea level [21]. The National Environment Management Authority, reports Moroto and Kotido districts in Karamoja region as semi-arid with distinct wet and dry seasons [7].
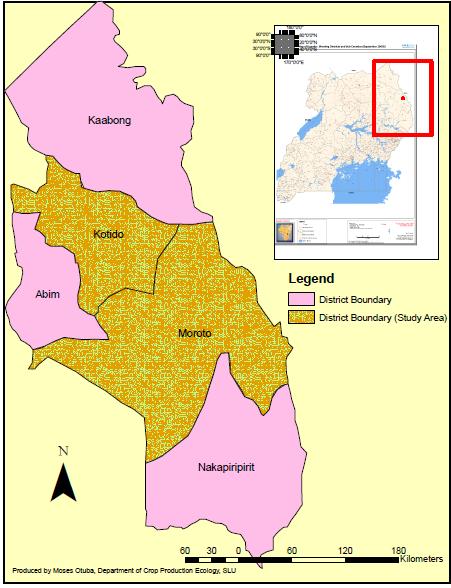 | Figure 1. Location of Kotido and Moroto districts in Uganda |
2.1. Measurements
- The shoot heights and leaf length of the two acacia species in the soil A and B substrates were measured before and after treatment of the seedlings with N fertilizer. Seedling heights were measured from the soil surface in the pots to the end of the shoot while the length of three leaves at bottom of each plant were measured from axillary bud by use of a tape measure.Leaf N concentration was determined from 80 acacia samples i.e 16 samples analyzed after first phase of seedling harvesting and then 64 after second phase using a Kjeldahl method [14] at the Department of Agricultural Production, Makerere University, Kampala in Uganda .
2.2. Statistical Data Analysis
- The functional approach to plant growth analysis [20] was used to compare seedling growth rates in various species and treatment combinations, by means of relative growth rates. Thus, the relative height growth RGRHT, and relative leaf length RGRLL were computed according to the following;
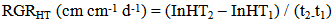 | (1) |
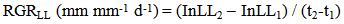 | (2) |
3. Results and Discussion
3.1. Growth Rate
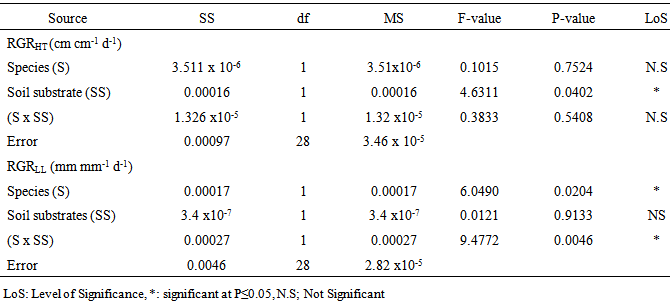
- Analysis of variance of data on growth rate showed that there was a significant effect (P≤0.05 ) of soil substrate on relative height growth (RGRHT), as well as significant species (S) effect and species-soil (S x SN) interaction on relative leaf length (RGRLL ) between the two acacia species (Table 1).
|
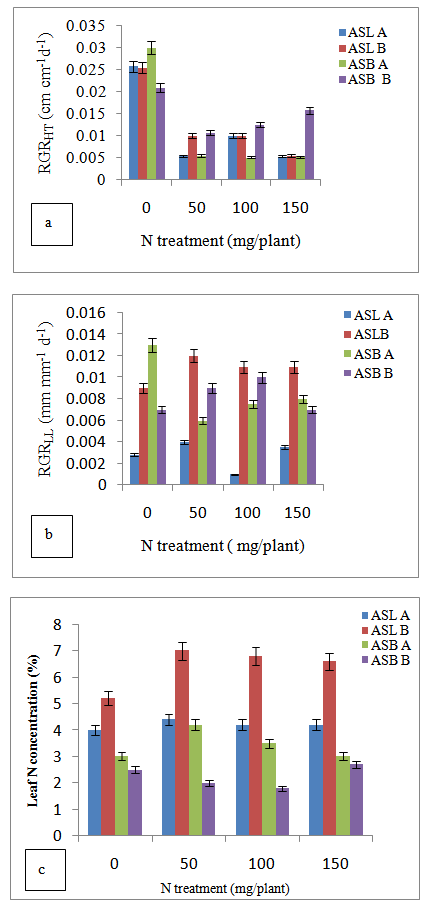 | Figure 2. (a) The influence of nitrogen (N) fertilizer on mean relative growth height (RGRHT) A. senegal (ASL) and A.sieberiana (ASB) seedlings in soil A and B, (b) A comparison of height of A.sieberiana seedlings in the pots treated with N fertilizer with those of N0 treatments in greenhouse. (c) The influence of nitrogen (N) fertilizer on mean relative leaf length (RGRLL) of A.senegal (ASL) and A.sieberiana (ASB) seedlings in soil A and B) |
3.2. Leaf N Concentration and Growth Traits
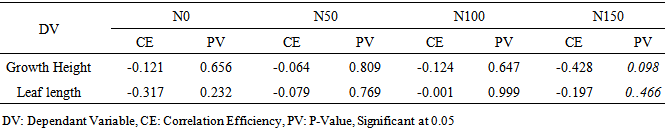
- The mean leaf N concentration of A. senegal and A. sieberiana in this study varied depending on the soil substrates and levels of the N treatments (Figure c). At no fertilization, A. senegal seedlings had higher leaf N concentration than A. sieberiana (Figure 2c). The seedlings of A. senegal in the soil substrates A and B treated with N fertilizers had higher leaf N concentration compared to those in soils with N0 treatments (Figure 2c). In all three N treatments (N50, N100 and N150), A. senegal had a higher leaf N concentration (7.1%, 6.6%, 6.3%) in soil B respectively compared to the N0 treatments.On the other hand, A. sieberiana had a lower leaf N concentration (1.9%, 1.6%) in soil B treated with N50 and N100 respectively over the N0 treatments (Figure 2c).Analysis of correlation of data on the growth traits indicated that there was no significant effect (P≤0.05) of the leaf N concentration on the growth height and leaf length of the two acacia species in all treatments (Table 2).There was a negative correlation between the leaf N concentration and growth traits, i.e. growth height and leaf length of the two acacia species in all treatments (Table 2). The growth height of the two acacia species had a higher negative correlation coefficient (-0.43) in soil A and B treated with N150 mg / plant after final harvest compared to other treatments (Table 2). However, the leaf length of the two acacia species had a lower negative correlation coefficient (-0.001) in soil A and B treated with N100 compared to other treatments (Table 2).
|
4. Conclusions
- Unfertilized soil substrates increased the growth rate of A. senegal and A. sieberiana seedlings more than the N fertilizer treatments. The two soil substrates affected the growth traits of the acacia species.The leaf N concentration also caused no impact on the growth traits of the two acacia species in all treatment.
ACKNOWLEDGEMENTS
- I would like to thank Swedish Institute, Department of Crop Production Ecology Swedish University of Agricultural Sciences and National Agricultural Research Organization for funding this study.
References
| [1] | Albert, V, Subramanian, A, Rangarajan, K and Pandy, R,M. 2011. Agreement of two different laboratory methods to measure electrolytes. Journal of Laboratory physicians,3(2), 104-109. |
| [2] | Alva, A.K., Paramasivam, S., Fares, A., Obreza, A., Schumann, A.W. 2006. Nitrogen best management practice for citrus trees II. Nitrogen fate transport and components of N budget. Scientia Horticulture 109: 223-233. |
| [3] | Bado, B.V, Sedogo, M.P, Lompo, F. 2004. Long term effects of mineral fertilizers, phosphate rock, dolomite and manure on the characteristics of an ultisol and maize yield in Burkina Faso. In: Bationo A (eds). Managing nutrient cycles to sustain soil fertility in Sub-saharan Africa.P.608. |
| [4] | Boussadia, O., Steppe, K., Zgallai., H, Ben El Hadj, S., Braham, M., Lemeur, R., Van Labeke, M.C. 2010. Effects of nitrogen deficiency on leaf photosynthesis, carbohydrate status and biomass production in two olive cultivars ‘Meski’ and ‘Koroneiki’. Scientia Horticulture, 123: 336-342. |
| [5] | Cakmak, I. 2002. Plant nutrition research: Priorities to meet human needs for food in sustainable ways. Plant and Soil, 247: 3-24. |
| [6] | Department of Meteorology Uganda. 2002. Increase Production with Meteorological Information, Agro Meteorological Bulletin vol.11, Issue 7. |
| [7] | Egadu, S. P., Mucunguzi, P., & Obua, J. 2007. The population of Acacia tree species producing gum arabic in the Karamoja region, Uganda. African Journal of Ecology, 45: 236-241. |
| [8] | Evans, E. 2000. A gardener’s Guide to Fertilising Trees and shrubs, NC State University. |
| [9] | Fernandez-Escobar. R, Benlloch. M, Herrera. E, Garcia-Novelo. J.M. 2004. Effects of traditional and slow-release N fertilizers on growth and Olive –nursery plants and N losses by leaching. Sci. Hort 101: 39-49. |
| [10] | Gradé, J. T., Tabuti, J. R. S. & Van Damme, P. 2009. Ethnoveterinary knowledge in pastoral Karamoja, Uganda. Journal of Ethnopharmacology, 122: 273-293. |
| [11] | Gruhn, P., Goletti, F., & Yudelman, M. 2000. Integrated nutrient management, soil fertility, and sustainable agriculture: current issues and future challenges. Food, Agriculture, and the Environment Discussion Paper 32, International Food Policy Research Institute, Washington, D.C. |
| [12] | He, Z.L., Calvert, D.V., Alva, A. K., Li, Y.C. 2000. Management of nutrients in citrus production systems in Florida. An overview. Soil Crop Sci. Fla. Proc. 59: 2-10. |
| [13] | Jacobs, D.F., & Timmer, V.R. 2005. Fertilizer induced changes in rhizosphere electrical conductivity: Relation to forest tree seedling root system growth and function. New For. 30:147–166. |
| [14] | Janssen, E, 2003. Determination of total phosphorus, total nitrogen and nitrogen fractions.Desk Study 16, Project Horizontal – Inorg. |
| [15] | Mehari, A.2005. Growth suitability of some tree species selected for planting in adverse environment in Eritrea and Ethiopia. Diss Uppsala; Swedish University of Agricultural Sciences. |
| [16] | Mindy, L., Bumgarner, K. Francis Salifu, and Douglass F. Jacobs. 2008. Subirrigation of Quercus rubra Seedlings: Nursery Stock Quality, Media Chemistry, and Early Field Performance HORTSCIENCE 43(7): 2179–2185. |
| [17] | Mugwe, J. Mugendi, Okobia, B, Tuwei, P, O’Neill, M. 2004. Soil conservation and fertility improvement using leguminous shrubs in central highlands of Kenya: NARFP case study. In: Bationo A (eds). Managing nutrient cycles to sustain soil fertility in Sub-saharan Africa.P.608. |
| [18] | Ndema, N.E, Etame, J, Taffouo, V.D and Bilong, P. 2010. Effects of some physical and chemical characteristics of soil on productivity and yield of Cowpea (Vigna unguicalata L.Walp) in Coastal region (Cameroon). African Journal of Environmental Science and Technology 4(3). pp108-114. |
| [19] | Raddad, E. Y., Luukkanen, O., Salih, A. A., Kaarakka, V., & Elfadl, M. A. 2006. Productivity and nutrient cycling in young Acacia senegal farming systems on Vertisol in the Blue Nile region, Sudan. Agroforestry Systems, 68: 193-207. |
| [20] | Rufat, J and Dejong, T.M., 2001. Estimating seasonal nitrogen dynamics in Peach trees in response to nitrogen availability. Tree physiology 21, 1133-1140. |
| [21] | Tessmer O.L, Jiao, Y, Cruz J.A, Kramer, D.M and Jin Chen. 2013. Functional approach to high-throughput plant growth analysis.BMC Systems Biology 2013; 7 (suppl6):S17. |
| [22] | Wekesa, C., Makenzi, P. M., Chikamai, B. N., Luvanda, A. M. & Muga, M. O. 2010. Traditional ecological knowledge associated with Acacia senegal (Gum arabic tree) management and gum arabic production in northern Kenya. International Forestry Review, 12: 240-246. |
| [23] | Wilson, J., & Rowland. 2001. Land and agriculture in Karamoja, Funded by European Union, Leeds. |
| [24] | Xu, X and Timmer, V.R. 1998. Biomass and nutrient dynamic of Chinese fir seedlings under conventional and experiment fertilization regimes. Plant and soil, 203: 313-322. |