-
Paper Information
- Next Paper
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2014; 4(5): 343-350
doi:10.5923/j.ijaf.20140405.01
Using Trichoderma viride for Optimization of Vermicomposting Processes to Improve the Quality of Vermicompost and Prolong the Storage Period
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLNatalia N. Tereshchenko, Alla B. Bubina, Tatiana V. Yunusova
Siberian Institute of Agriculture and Peat, Siberian Branch, Russian Academy of Agricultural Sciences, Tomsk, 634050 Russia
Correspondence to: Natalia N. Tereshchenko, Siberian Institute of Agriculture and Peat, Siberian Branch, Russian Academy of Agricultural Sciences, Tomsk, 634050 Russia.
| Email: |  |
Copyright © 2014 Scientific & Academic Publishing. All Rights Reserved.
The performed study demonstrates that a preliminary fermentation of organic substrate with Trichoderma is promising, since this allows the vermicultivation time to be reduced and the yield and quality of vermicompost to be elevated. Vermicompost obtained after pre-fermentation of organic substrate with Trichoderma has the maximal indicators of growth-stimulating activity and the highest level of fungistatic properties in relation to root rot pathogens. This variant of vermicompost maintains the highest level of biological activity even after 8 months of storage. During the storage period growth-stimulating activity of vermicompost changes cyclically. The level of growth promoting activity of vermicompost is greatly influenced not only by period of storage, but also by the stage of organic matter transformation in vermicompost at the end of the process of vermicultivation. Excessive prolongation of vermicultivation causes a decrease in its biological activity.
Keywords: Vermicompost, Vermicultivation, Trichoderma viride, Introduction, Growth-stimulating activity, Fungistatic activity, Storage period
Cite this paper: Natalia N. Tereshchenko, Alla B. Bubina, Tatiana V. Yunusova, Using Trichoderma viride for Optimization of Vermicomposting Processes to Improve the Quality of Vermicompost and Prolong the Storage Period, International Journal of Agriculture and Forestry, Vol. 4 No. 5, 2014, pp. 343-350. doi: 10.5923/j.ijaf.20140405.01.
1. Introduction
- The efficiency of introduction of the active fungal strains belonging to the genus Trichoderma into the plant rhizosphere, as well as preplanting inoculation, has been demonstrated in numerous studies [1–3]. Despite the fact that preparations involving Trichoderma as the major component is a real alternative to synthetic agrichemicals as antagonists of plant soil diseases and growth stimulators, their commercial application presents certain difficulties. Application of such preparations does not always give stable results, especially under field conditions [4]. Ecological studies have shown that fungal strains belonging to the genus Trichoderma can not always be successfully introduced into the soil microbial community due to their low competitiveness and high demand for available sources of carbon. For this reason, their influence as biological control agents is limited to a few weeks after introduction [5].Besides some technological problems caused by short shelf life of the liquid form of the preparations based on Trichoderma frequently take place. Whereas the use of the solid preparation form often causes the problem of autoinhibition of conidia germination. For this reason, there is a need for a careful calculation of the optimal dose for each particular strain [6]. Therefore, development of multifunctional biological preparations stable in their efficiency, including the preparations involving Trichoderma, is among the most important tasks in the system of developing organic crop farming. In order to maintain a stable population of Trichoderma in the soil during the growing season, some authors propose to perform the application of Trichoderma simultaneously with the introduction of various organic substrates: wheat bran, peat moss, pine bark, compost from cow manure and even waste substrates after oyster culturing [7, 8].In our opinion, a joint application of Trichoderma and vermicompost is promising technological method to raise Trichoderma survival in the soil and fungal colonization of the plant rhizosphere. On the other hand, since the quality of vermicompost depends on the content and activity of the agronomically valuable microflora, vermicompost enrichment for the active microbial strains responsible for fungistatic, growth-stimulating, and other beneficial properties can have a positive effect on the efficiency of its use. It is also known that many species of the genus Trichoderma are able to accelerate the composting rate of organic substrates due to their ability to transform the lignin–cellulose complex with their cellulolytic enzymes, which, in turn, enriches the final product for nutrients [9].In accordance with the above arguments the goal of this work was to improve the technology for producing vermicompost by preliminary fermentation of organic substrate using the fungus Trichoderma viride with the aim to reduce the time required for vermicultivation and to elevate the growth-stimulating and fungistatic activity of vermicompost.
2. Experimental
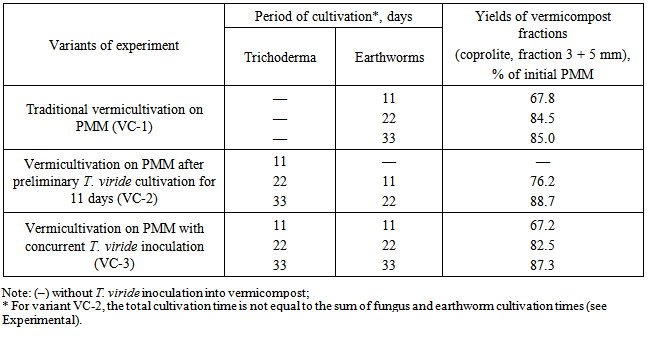
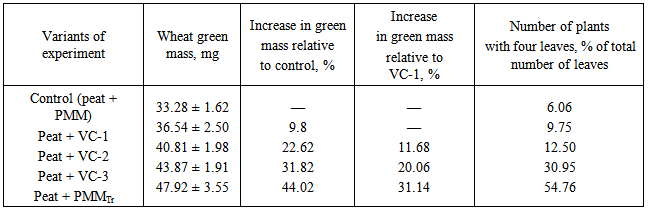
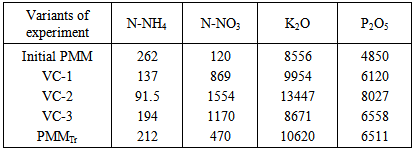
- Objects of study were the earthworm Eisenia andrei and a pure culture of the saprophytic fungus Trichoderma viride, isolated by the authors from pine wood. The growth-stimulating activity of vermicompost was determined by biological tests and vegetation experiments with the wheat cultivar Tulunskaya 12. The earthworms were cultivated in a peat–manure mixture (PMM) composed of fen peat from the Kandinskoe deposit and cattle litter manure at a ratio of 1: 4 (wt/wt) with 40 mature earthworm individuals (30 g) added per 1 kg substrate. The moisture content in the substrate was maintained at a level of 80% and temperature was maintained in the range of 20–25°C.The model experiment comprised several variants, namely, (1) traditional vermicultivation on PMM for 33 days (VC-1); (2) vermicultivation on PMM after preliminary solid-phase cultivation of the fungus T. viride for 11 days, with the period of vermicultivation per se of 22 days (VC-2); (3) vermicultivation on PMM for 33 days with concurrent introduction of the fungus T. viride into the substrate (VC-3); (4) PMM composting with introduction of the fungus without vermicultivation (PMMTr). During vermicultivation the efficiency of the initial substrate conversion into compost by earthworms was determined in all experimental variants. For this purpose, the fractions of coprolite were separated using 3- and 5-mm sieves to calculate the percent rate of each fraction.In the vegetation experiment, peat supplemented with 25% of PMM was used as a control. The experimental soil variants were composed of the peat supplemented with 25% of the corresponding vermicompost variants (VC-1, VC-2, or VC-3) or 25% of the PMMTr relative to the total weight of the prepared substrate. The experiment was performed in seven replicates using plastic containers (500 ml) with six wheat grains planted in each. The wheat was grown at a temperature of 25 ± 1°C in a phytochamber with daylight of 15 h. Biological activity of vermicompost was assessed according to the increase in wheat green mass (dried to a constant weight) expressed as percent of the control. Growth-stimulating activity was determined by biological testing of wheat seeds soaked for 5 min in an acid extract (pH 4–4.5) of vermicompost. Stimulatory effect was assessed according to the difference between the dry weights of green shoots and roots in the experimental and control (with distilled water) variants expressed as percent of the control.T. viride enrichment culture was produced on wheat grain preliminary boiled to softness in a small amount of water. The grain was placed into bottles; the bottles were stopped and twice sterilized in an autoclave at 1 atm for 30 min with an interval of 1 day. Then, the tubes with the grain inoculated with T. viride were kept in a thermostat at a temperature of 28–30°C until fungal mycelium completely covered the grain. The produced enrichment culture was introduced into the substrate in an amount of 2% of the PMM weight in the container. Since the fungal population on the PGA (potato–glucose agar) medium was reduced due to active growth of the colonies and competition for the substrate, the T. viride during vermicultivation was counted on MPA (meat–peptone agar) [10].Vermicomposts and initial organic substrate were chemically assayed for the following components: nitrate-nitrogen using phenol disulfonic acid (state standard GOST 26488-85), ammonium nitrogen using the Nessler reagent (GOST 26489-85), labile phosphorus according to Kirsanov (GOST 26207-91), and exchange potassium according to Kirsanov using lame photometry (GOST 26207-91). To assess the impact of the storage period on the vermicomposts quality (growth-stimulating and fungistatic activity) the model experiment with the cultivation of wheat seedlings in peat containing soil was conducted after 3 months of storage. All variants of vermicompost were stored at 5°C. In each experimental variant the soil was supplemented with 25% of previously obtained corresponding vermicompost variants (VC-1, VC-2, VC-3). As a control we used the soil supplemented with 25% of peat-manure mixture (PMM). To evaluate the level of fungistatic activity of vermicomposts the infectious load was artificially modeled in the experiment by adding mycelium and spores of Bipolaris sorokiniana into the soil as a part of the chopped agar medium Capek. The model experiment include the following variants: (1) Soil + VC-1; (2) Soil + VC-1 + Bipolaris sorokiniana; (3) Soil + VC-2; (4) Soil + VC-2 + Bipolaris sorokiniana; (5) Soil + VC-3; (6) Soil + VC-3 + Bipolaris sorokiniana. The experiment was performed in five replicates using plastic containers (500 ml) with six wheat grains planted in each. The wheat was grown at a temperature of 25 ± 1°C in a phytochamber with daylight of 15 h. Data were statistically processed using Statistica v. 5.5 and 6.0 software packages [11]. In the paper, the data are presented as a mean value with a confidenc entervals for variance and standard deviation calculated using Student’s test (p < 0.05). Data on wheat growth in the vegetation experiment were compared according to the nonparametric Mann–Whitney test (p < 0.05).
3. Results and Discussion
- Determination of the efficiency of organic substrate conversion by earthworms have demonstrated that preliminary fermentation of the substrate with T. viride (VC-2) allowed the yield of vermicompost to be elevated by 8.5% as compared with traditional vermicultivation (VC-1) as early as day 11 (Table 1).
|
|
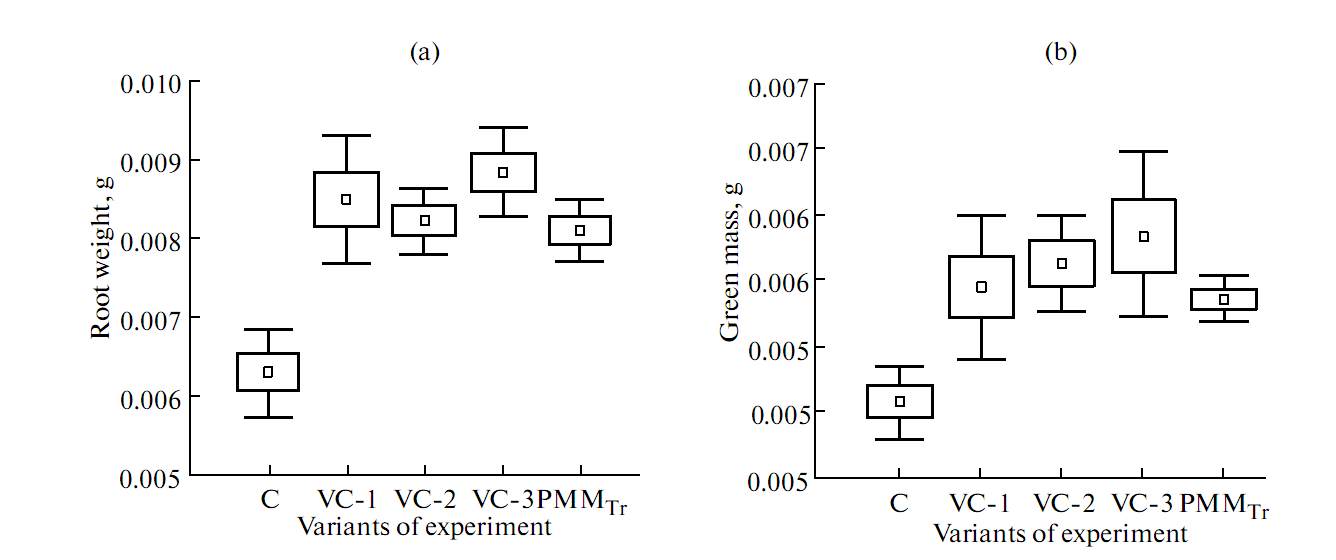 | Figure 1. Efficiency of acid extracts derived from the vermicompost variants (VC-1, VC-2, VC-3) and compost produced by T. viride fermentation (PMMTr) on the root weight (a) and green mass (b) of wheat seedlings |
|
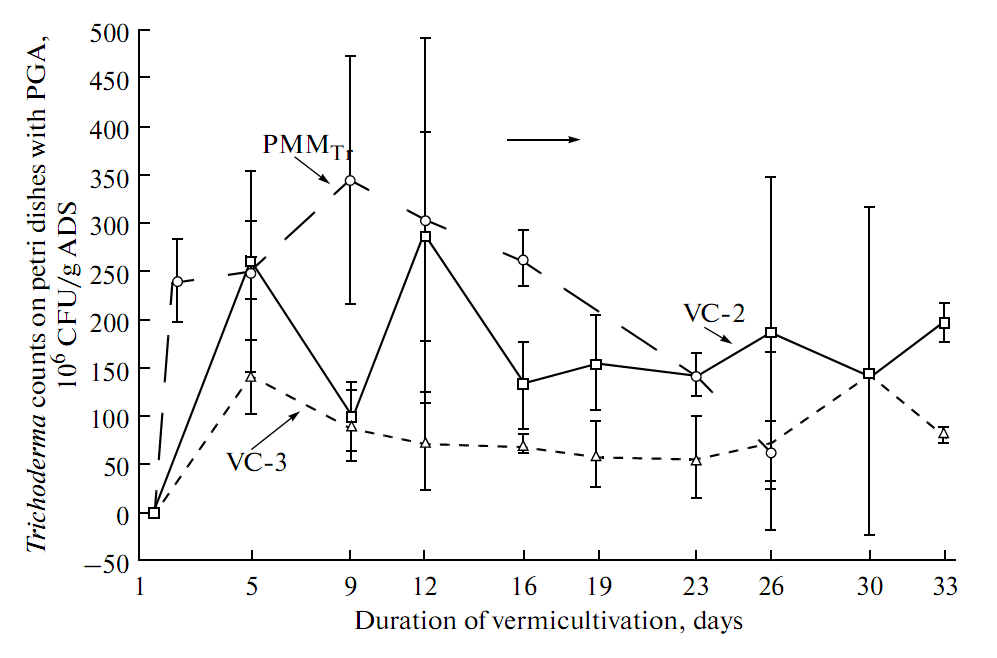 | Figure 2. Dynamics of T. viride population in the vermicompost variants (VC-1, VC-2, VC-3) and compost produced by fermentation of organic substrate with T. viride (PMMTr) |
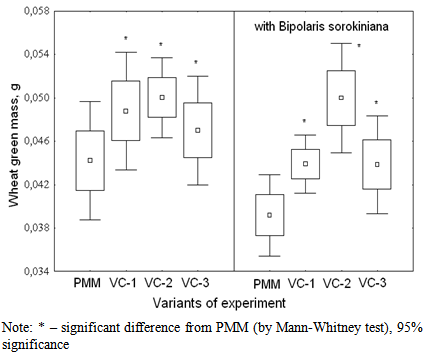 | Figure 3. Effect of vermicomposts (VC-1, VC-2, VC-3) on the formation of wheat green mass after 3 months of storage |
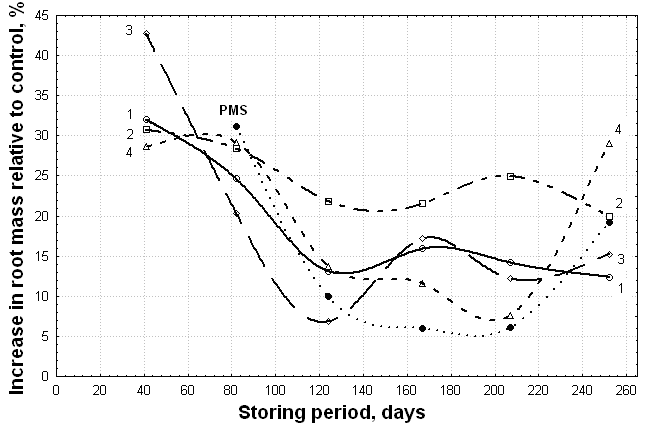 | Figure 4. Dynamics of the activity of acid extracts derived from vermicomposts (1 –VC-1, 2 – VC-2, 3 – VC-3) and compost produced by T. viride fermentation (PMMTr) in relation to the roots of wheat seedlings compared with the activity of acid extracts derived from initial peat-manure substrate (PMS) within 260 days of storage |
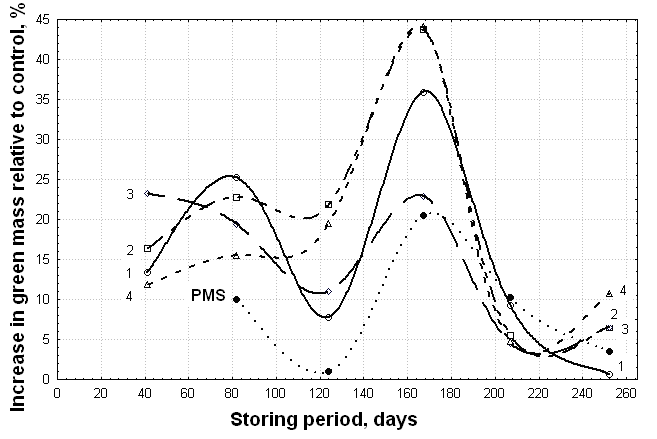 | Figure 5. Dynamics of the activity of acid extracts derived from vermicomposts (1 –VC-1, 2 – VC-2, 3 – VC-3) and compost produced by pre-fermentation of substrate with T. viride (PMMTr) in relation to the wheat green mass compared with the activity of acid extracts derived from initial peat-manure substrate (PMS) within 260 days of storage |
4. Conclusions
- Thus, a preliminary composting of peat–manure mixture using Trichoderma viride is a promising technological method for accelerating the conversion of organic substrate during vermicultivation, which enhances an increase in production efficiency. An increase in the rate of organic substrate conversion does not decrease the quality of the produced vermicompost but, on the contrary, enhances its enrichment for nutrients and increase in its growth-stimulating activity.High level of fungistatic activity of vermicompost persists for at least 3 months from the beginning of the storage period. The maximum level of resistance of wheat seedlings against root rot (Helminthosporium) has VC-2 obtained after preliminary fermentation substrate by Trichoderma.Growth-stimulating activity of vermicompost remains high, at least for 260 days from the beginning of the storage period. During the storage period growth-stimulating activity of vermicompost changes cyclically.The level of growth promoting activity of vermicompost is greatly influenced not only by period of storage, but also by the stage of organic matter transformation in vermicompost at the end of the process of vermicultivation. Definition and strict adherence to the optimal period of vermicultivation ultimately provides a production of vermicompost characterized by high level of growth-stimulating activity and high stability during period of storage. Excessive prolongation of vermicultivation causes a decrease in its biological activity.