-
Paper Information
- Next Paper
- Previous Paper
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Agriculture and Forestry
p-ISSN: 2165-882X e-ISSN: 2165-8846
2012; 2(1): 58-62
doi:10.5923/j.ijaf.20120201.10
Combining Ability Analysis of Plant Height and Yield Components in Spring Type of Rapeseed Varieties (Brassica napus L.) Using Line × Tester Analysis
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLValiollah Rameeh
Agricultural and Natural Resources Research Center of Mazandran, Sari, Iran
Correspondence to: Valiollah Rameeh, Agricultural and Natural Resources Research Center of Mazandran, Sari, Iran.
| Email: |  |
Copyright © 2012 Scientific & Academic Publishing. All Rights Reserved.
Line ×tester of six lines and two testers of spring rapeseed (Brassica napus L.) cultivars were used to estimate general combining ability (GCA), specific combining ability (SCA) effects, high parent heterosis and narrow-sense heritability estimate for plant height, yield components and seed yield. Significant variance of line x tester for pods per plant and seed yield, indicating non additive genetic effects have important role for controlling these traits. Significant mean squares of parents vs crosses which are indicating significant average heterosis were also significant for all the traits except seeds per pod. High narrow-sense heritability estimates for all the traits except seeds per pod exhibited the prime importance of additive genetic effects for these traits except seeds per pod. Most of the crosses with negative SCA effect for plant height had at least one parent with significant negative or negative GCA effect for this trait. For most of the traits except pods per plant, the efficiency of high parent heterosis effect was more than SCA effect for determining superior cross combinations.
Keywords: Additive, GCA, Heritability, Line × Tester, Yield Components
Cite this paper: Valiollah Rameeh, Combining Ability Analysis of Plant Height and Yield Components in Spring Type of Rapeseed Varieties (Brassica napus L.) Using Line × Tester Analysis, International Journal of Agriculture and Forestry, Vol. 2 No. 1, 2012, pp. 58-62. doi: 10.5923/j.ijaf.20120201.10.
Article Outline
1. Introduction
- Rapeseed (B. napus L.) has important role in oilseed production in Iran and other parts of the world because of its wide adaptation to different diverse climatic conditions. Hence, it has become necessary to enhance the present production by developing superior varieties of rapeseed. Improvement of superior varieties could be done by reshuffling the genes through hybridization from proper parents. Moreover, it is also necessary to know about the nature and magnitude of gene action responsible for controlling the inheritance of various yield attributes along with combining ability of the parents and their cross combinations in order to make use of them in further crop improvement program. Seed yield of canola is a quantitative trait, which is largely influenced by the different environmental effects and hence in most of the cases it has low heritability (Habekotte, 1997; Diepenbrock, 2000; Rameeh, 2010). In rapeseed breeding program for hybrid and open pollinated varieties, general and specific combining ability effects (GCA and SCA) are important indicators of the potential of inbred lines in hybrid combinations. The line×tester analysis is one of the efficient methods of evaluating large number of inbred as well as providing information on the relative importance of GCA and SCA effects for interpreting the genetic basis of important plant traits (Singh and Chaudhury, 1977). By using this scheme, and other genetic designs like diallel analysis significant GCA and SCA effects of phenological traits, seed yield and other yield associated traits were reported in rapeseed (Shen et al., 2002; Nassimi, et al., 2006b; Wang et al., 2007). In earlier studies on spring cultivars of rapeseed (Huang et al., 2010; Rameeh, 2011) were stressed the important role of GCA and SCA effects for yield components.Mid parents and high parent heterosis (heterobeltiosis) have extensively been explored and utilized for boosting various quantity and quality traits in rapeseed (Nassimi, et al., 2006a; Chapi et al., 2008). With sufficient level of heterosis, commercial production of hybrid varieties would be justified (Nassimi, et al., 2006a). For seed yield in spring rapeseed hybrids, an average high parent heterosis of 30% with a range of 20–50% was observed, while for winter rapeseed hybrids an average high parent heterosis of 50% was reported, ranging from 20 to 80% as reviewed by McVetty (1995). The unset of flower initiation can have strong influence on flower, pod and seed number (Diepenbrock, 2000; Downey and Rimer, 1993; Yasari and Patwardhan, 2006). Habekotte, (1997) used a sensitivity analysis within a crop growth model to study options for increasing seed yield in winter oilseed rape. The most promising crop type for high seed yield combined late maturity with early flowering (Downey and Rimer, 1993). Heterosis is commercially exploited in rapeseed (Brassica napus L.) and its potential use has been demonstrated in turnip rape (B. rapa L.) and Indian mustard (B. juncea L) for most agronomic traits (Teklwold and Becker, 2005; Zhang and Zhu, 2006).Due to different genetic control of yield associated traits in various genetic materials, the objectives of the present study were therefore to identify general and specific combining abilities, heterosis and narrow-sense heritability estimates for plant height and yield associated traits in adapted rapeseed spring cultivars.
2. Materials and Methods
- Six spring rapeseed (B. napus L.) cultivars including Sarigol, RGS3006,RGS003, 19H, RW and Option500 as lines were crossed with two spring testers including R1 and R2 based on line x tester crossing method during 2005-06. Twelve F1s along with their parents were sown in a randomized complete block design with three replications at Biekol Agriculture Research Station, located in Neka, Iran (53˚, 13′ E longitude and 36˚ 43′ N latitude, 15 m above sea level) during winter 2006-07. Each plot was consisted of four rows 5 m long and 40 cm apart. The distance between plants on each row was 5 cm resulting in approximately 300 plants per plot, which were sufficient for F1 genetic analysis. The soil was classified as a deep loam soil (Typic Xerofluents, USDA classification) contained an average of 280 g clay kg-1, 560 g silt kg-1, 160 g sand kg-1, and 22.4 g organic matter kg-1 with a pH of 7.3. Soil samples were found to have 45 kg ha-1 (mineral N in the upper 30-cm profile). Fertilizers were applied at the rates of 100: 50: 90 kg/ha of N: P: K, respectively. All the plant protection measures were adopted to make the crop free from insects. The data were recorded on ten randomly selected plants of each entry of each replication for plant height, pods per plant, seeds per pod and 1000-seed weight. Seed yield (adjusted to kg/ha) was recorded based on two middle rows of each plot.Data for the genotypes were subjected to line x tester analysis (Singh and Chaudhury 1977) to estimate general combining ability (GCA) and specific combining ability (SCA). A t-test was used to test whether the GCA and SCA effects were different from 0. For each hybrid and each studied trait, the difference between hybrid and the mean of high parent was computed separately. A least significant difference (LSD) was used to test whether these differences were different from 0.
3. Results and Discussion
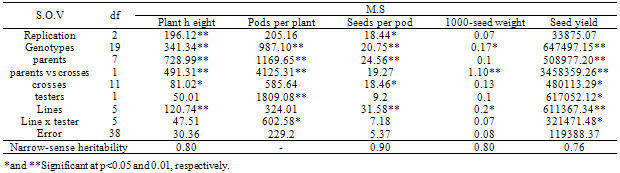
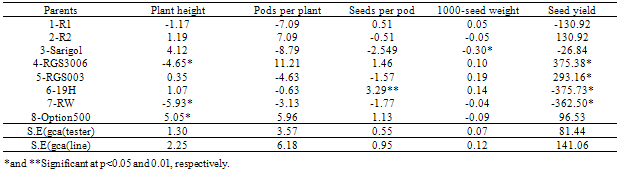
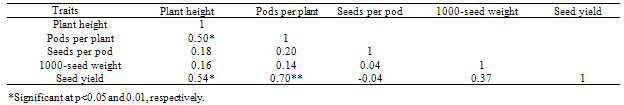
- Line x tester analysis of varianceSignificant mean square of the genotypes for the traits under study including plant height, pods per plant, seeds per pod, 1000-seed weight and seed yield indicating enough genetic variations for the genotypes and necessity of genetic analysis (Table 1). Significant mean squares of parents vs crosses which are indicating significant average heterosis were also significant for all the traits except seeds per pod. Significant mean squares of testers were detected for pods per plant and seed yield and significant mean squares of lines were determined for all the traits except pods per plant. High narrow-sense heritability estimates for all the traits except seeds per pod exhibited the prime importance of additive genetic effects for these traits except seeds per pod. Significant mean squares of line x tester for pods per plant and seed yield, indicating non additive genetic effects have important role for controlling these traits. Significant GCA and SCA effects of yield components were reported for spring (Rameeh, 2010) and winter (Huang et al., 2010; Sabaghnia et al., 2010) types of rapeseed. Although these researchers were noted the more importance of non additive genetic effects for yield associated traits but it can be different based on genotypes under study and in this study additive genetic effect was more important for most of yield associated traits.General combining ability effects of the parentsSignificant negative GCA effect of plant height was displayed for RGS003 and RW and due to importance of dwarf for ideotype breeding of rapeseed, these parents are favorite combiners for improving this trait (Table 2). Regards to important of non additive genetic effects for pods per plant, non of parents had significant GCA effects for this trait. 19H had significant positive GCA effect for seeds per pod. RGS003 and 19H with positive GCA effect for 1000-seed yield were considered as good combiners. Significant positive GCA effect of seed yield was detected for RGS3006 and RGS003, therefore these parents were considered as desirable combiners for seed yield improving. Many studies have been made in rapeseed on the inheritance of economic characters, especially seed yield and oil content, but the results were variable. In their materials, Brandle and McVetty (1990) reported that GCA accounted for 88 and 44% of hybrids sum of squares for seed yield, respectively. Shen et al. (2002) found that GCA accounted for 76.65% of the hybrid sum of squares for seed yield. Significant negative GCA effects were reported for yield components (Huang et al., 2010; Rameeh, 2010; Sabaghnia et al., 2010). Significant positive correlation was determined between pods per plant and seed yield, therefore this trait can be considered as suitable selection criterion for seed yield improving (Table 3).Specific combining ability effects of the crossesThe crosses including Sarigol x R1, RW x R1, Option500 x R1, RGS003 x R2, RGS3006 x R2 and 19H x R2 with negative SCA effects for plant height were detected as good cross combinations for decreasing this trait (Table 4). Most of the crosses with negative SCA effect for plant height had at least one parent with significant negative or negative GCA effect for this trait. Nassimi et al. (2006b) noted that in Brassica (grown for seed yield), taller plants are susceptible to lodging. Medium-or short-statured plants are therefore desirable, and accordingly negative GCA and SCA values are sought for plant height. The same researchers have found that out of 28 crosses, 13 showed negative SCA effects for plant height. Out of 12 crosses, 6 crosses had significant SCA effects of pods per plant. The crosses including RGS3006 x R1, Option x R1, Sarigol x R2 and RGS003 x R2 with significant positive SCA effects for pods per plant were good combinations for increasing this trait and due to significant positive correlation of this trait with seed yield, selection of these crosses will improve the seed yield. Non of crosses had significant SCA effect for seeds per pod, 1000-seed weight and seed yield. The crosses including 19H x R1, RGS3006 x R2 and Option x R2 had positive SCA effect for seeds per pod. The crosses viz. Sarigol x R1, RW x R1 and RGS3006 x R2 with positive SCA effect for 1000-seed weight were considered as suitable combinations for improving this trait. The combinations including RW x R1, Sarigol x R2 and 19H x R2 with positive SCA effect for seed yield were good crosses for increasing seed yield. Similarly in earlier study (Nassimi et al. 2006b) was reported significant SCA effects for plant height, the number of pods per main raceme, the number of primary branches per plant, days to 50% flowering and physiological maturity
|
|
|
|
|
4. High Parent Heterosis
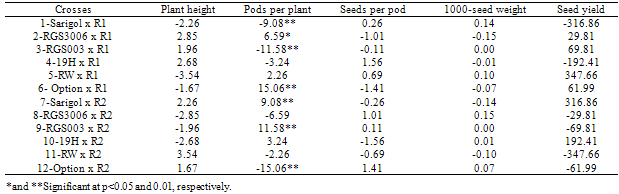
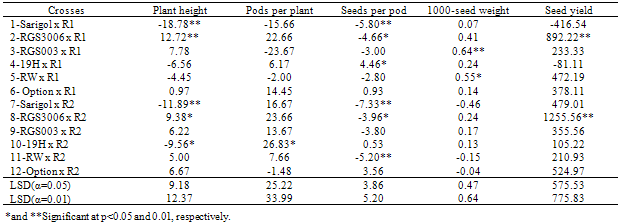
- The result of high parent heterosis effects of crosses for all the traits is presented in Table 5. Out of 12 crosses, 4 crosses had significant high parent heterosis effects of plant height. The crosses including Sarigol x R1, Sarigol x R2 and 19H x R2 with significant negative high parent heterosis for plant height were suitable combinations for deceasing this trait. Although most of the crosses had significant SCA effect for pods per plant, only one of combination had significant high parent heterosis effect for this trait. 19H x R2 with significant positive high parent heterosis effect for pods per plant was good combination for improving this trait. Although non of crosses had significant SCA effect for seeds per pod, six crosses had significant high parent heterosis effects for this trait. 19H x R1 with significant positive high parent heterosis effect for seeds per pod was good combination for increasing this trait and both of parents of this combination had positive GCA effect for seeds per pod. RGS003 x R1 and RW x R1 with significant positive high parent heterosis effects for 1000-seed weight were considered as suitable combinations for improving this trait. Out of 12 crosses, 2 crosses had significant high parent heterosis effects of seed yield. For seed yield in spring rapeseed hybrids, an average high parent heterosis of 30% with a range of 20–50% was observed, while for winter rapeseed hybrids an average high parent heterosis of 50% was reported, ranging from 20 to 80% as reviewed by McVetty (1995).In general high narrow-sense heritability estimates for all the traits except seeds per pod exhibited the prime importance of additive genetic effects for these traits except seeds per pod. Significant variance of line x tester for pods per plant and seed yield, indicating non additive genetic effects have important role for controlling these traits. Although most of the crosses had significant SCA effects for pods per plant, but one cross had significant high parent heterosis effect for this trait, therefore the efficiency of SCA effect is more important than heterosis effect for detecting superior combinations for pods per plant. For other studied traits, the efficiency of high parent heterosis effect is more than SCA effect for determining superior cross combinations.
ACKNOWLEDGEMENTS
- The author wish to thanks from Agricultural and Natural Resources Research Center of Mazandaran and Seed and Plant Improvement Institute (SPII) for providing genetic materials and facility for conducting the experiment.