-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
American Journal of Chemistry
p-ISSN: 2165-8749 e-ISSN: 2165-8781
2021; 11(4): 59-65
doi:10.5923/j.chemistry.20211104.01
Received: Aug. 27, 2021; Accepted: Sep. 10, 2021; Published: Sep. 26, 2021

Insecticidal Drimane Sesquiterpenes from Warburgia ugandensis against Maize Pests
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLSylvia A. Opiyo
Department of Physical and Biological Sciences, Murang’a University, Murang’a, Kenya
Correspondence to: Sylvia A. Opiyo , Department of Physical and Biological Sciences, Murang’a University, Murang’a, Kenya.
| Email: |  |
Copyright © 2021 The Author(s). Published by Scientific & Academic Publishing.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Sitophilus zeamais and Prostephanus truncatus, the destructive pests of maize in Africa, cause extensive tunneling in maize grain leading to conversion of maize grain into flour within a very short time. Small-scale farmers are often forced to sell maize shortly after harvest to minimize losses during storage, thereby attract low prices and compromising food security. Most available pesticides have adverse effects on environment and humans. The aim of this study was to evaluate efficacy of extracts and compounds of Warburgia ugandensis in controlling of S. zeamais and P. truncatus. Chromatographic fractionation of extracts from W. ugandensis afforded 7α-acetylugandensolide (1), ugandensolide (2), polygodial (3), warbuganal (4), ugandensidial (5), mukaadial (6) and muzigadial (7). The structures of the compounds were determined using spectroscopic and physical methods. n-Hexane extract was the most repellent among the extracts while polygodial (3) was the most repellent compound against the two pests. n-Hexane and ethyl acetate extracts caused 76.3-78.3% and 71.1-75.0% deaths of the insects respectively. Polygodial (3) and warbuganal (4) caused 64.3-70.0 and 61.7-65.0% deaths respectively. n-Hexane and ethyl acetate extracts, polygodial (3) and ugandensolide (2) significantly inhibited the emergence of the insects. The findings from this study show that extracts from W. ugandensis are effective in controlling maize insect pests.
Keywords: Zea may, Insect pests, Repellence, Mortality, Emergence inhibition
Cite this paper: Sylvia A. Opiyo , Insecticidal Drimane Sesquiterpenes from Warburgia ugandensis against Maize Pests , American Journal of Chemistry, Vol. 11 No. 4, 2021, pp. 59-65. doi: 10.5923/j.chemistry.20211104.01.
Article Outline
1. Introduction
- Maize (Zea mays) is one of the major cereal grains grown in abundance in sub-Saharan Africa during the raining season. Maize is ranked fourth most edible grain after sorghum, millet and rice [1]. The crop accounted for 19.5% calorie being the world’s highest supplier of calorie for body growth, followed by rice (16.5%) and wheat which accounted for 15.0% [1]. Peasant farmers produce huge tons of maize annually which is usually more than enough for sale in the markets. This has resulted into wastage due to inadequate storage structures and insect pest which cause significant losses. Maize weevil (Sitophilus zeamais Motchulsky) and larger grain borer (Prostephanus truncatus Horn) are the most important pests of maize. The extensive tunneling in maize grain by pests allows the pests to convert maize grain into flour within a very short time [2]. Small-scale farmers are often forced to sell maize shortly after harvest to minimize losses during storage, thereby attract low prices and compromising food security. Chemical control is the most commonly used and most effective method at the farm level. However, some of the synthetic chemicals have adverse effects on environment and humans [3]. This underscores the need for affordable alternatives that are convenient to use and are environmentally friendly. Plant extracts contain secondary metabolites some of which inhibit the growth of pests and pathogenic microorganism [4-18]. The use of botanical for pests and disease control is preferred because they are safe and non-toxic to humans [19-24]. In addition, chances of pests and pathogens developing resistance to botanical pesticides are highly unlikely [25]. Warbugia ugandensis (Canellaceae) is used as a remedy for stomachache, constipation, toothache, malaria, sexually transmitted diseases, diarrhoea, cough and internal wounds/ulcers in ethno- medicine [14]. Plants belonging to the genus Warburgia are characterized by the presence of drimane sesquiterpenes which have been reported to exhibit antibacterial, antifungal, insect antifeedant, insecticidal and molluscicidal activities [26-28]. The present study reports the efficacy of extracts and compounds of W. ugandensis in controlling of S. zeamais and P. truncatus infestation in stored maize.
2. Materials and Methods
2.1. General Experimental Procedure
- Melting points were determined on a Gallenkamp (Loughborough, UK) melting point apparatus and are uncorrected. The UV spectra were run on Pye Unicam SP8-150 UV-vis pectrophotometer (Cambridge, UK) using acetonitrile. IR data were recorded on a PerkinElmer FTIR 600 series spectrophotometer (Waltham, MA, USA) as KBr pellet. 1H and 13C NMR data were measured in CDCl3 and CDCl3-DMSO-d6 on a Bruker NMR Ultrashield TM (Darmstadt, Germany) operating at 500 and 125 MHz, respectively. The MS data were obtained on a Varian MAT 8200A instrument (Bremen, Germany).
2.2. Plant Materials
- Warburgia ugandensis stem bark was collected form Nakuru-Gilgil Highway near St. Mary's Hospital (latitude 0° 24' 42.49'' S and longitude 36° 15' 10.59'' E) and plant specimen (2014/5/SAO/CHEMMK) was identified at the Kenya National Museum Herbarium after comparison with authentic samples. The materials were air dried at room temperature under the shade until crispy. The dried materials were pulverized and sieved through a 0.5 mm size mesh.
2.3. Extraction and Isolation of Compounds
- Two kg of powdered W. ugandensis stem bark was cold extracted with organic solvents of varying polarities (n-hexane, ethyl acetate and methanol) sequentially by soaking in the solvents for seven days with occasional shaking. The mixture was filtered and concentrated using a rotary evaporator at reduced pressure to yield 20.2, 58.6 and 97.8 g of n-hexane, ethyl acetate and methanol extracts, respectively. The resultant extracts were stored at 4°C for bioassays and phytochemical studies. Hexane and ethyl acetate extracts showed similar TLC profile and were combined for phytochemical isolation. The combined extract (50 g) was dissolved in a small amount of n-hexane - ethyl acetate mixture (1:1) and subjected to in silica gel for column chromatography using silica gel. Elution was done using n-hexane, n-hexane - ethyl acetate mixture, ethyl acetate and methanol to give 200 fractions (each 20 ml) whose compositions were monitored by TLC and those with similar profiles were combined to give seven pools labeled I -VII. Pool I, 3g, which was eluted with n-hexane did not show any major spot in TLC and was discarded. Pool II (7 g) was subjected to further column chromatography eluting with n-hexane: ethyl acetate (95:5, 9:1, 85:15 and 4:1) to give polygodial 30 mg and warbuganal 55 mg. Pool IV (5 g) on further fractionation with gradient n-hexane-ethyl acetate mixture (85:15, 4:1 and 7:3) gave polygodial 35 mg and ugandensolide 38 mg. Pool V (8 g) on further fractionation with n-hexane: ethyl acetate (4:1, 7:3 and 65:35) gave ugandensolide 24 mg, ugandensidial 42 mg and muzigadial 75 mg. Pool VI (9 g) gave ugandensidial 43 mg while Pool VII gave muzigadial 15 mg, 7α-acetylugandensolide 64 mg and mukaadial 72 mg.
2.4. Mass Rearing of Test Insects
- Adult insects were obtained from infested maize grains obtained from local market and a new generation was reared from the stock on dry pest susceptible maize grains [29]. Two hundred unsexed insects were introduced into a two-liter glass jars containing 400 g weevil susceptible maize grains [30]. The mouths of the jars were then covered with nylon mesh held in place with rubber bands and the jars left undisturbed for 35 days for oviposition. Thereafter, all adults were removed through sieving and each jar was left undisturbed for another 35 days. Emerging adult insects were collected and kept in separate jars according to their age. Adults that emerged on same day were considered of the same age [31].
2.5. Repellency Test
- The test was done according to (Mwangangi and Mutisya [29] with some modifications. Transparent plastic tubings, 13 cm long x 1.3 cm diameter were used as test cylinders. Each test cylinder was plugged at one end with cotton ball containing crude extracts and compound isolated from the stem bark of W. ugandensis while the other end was plugged with clean cotton ball which served as control. Actellic dust was used as a positive control. Ten-three-day old unsexed test insects were introduced at the middle of each test cylinder through a hole at the middle portion of the cylinder (0.0 cm) and let to move in any direction of their choice with scoring of distance moved measured in cm using a ruler. The score time was 2 and 24 hours after exposure time and all tests were done in triplicates.
2.6. Adult Mortality Test
- Contact toxicity assay was done according to Ileke and Oni [32] with some modifications. Toxicity of the crude extracts and isolated compounds were tested against adult insects. The test samples were mixed with talc thoroughly and the dust was admixed with 20 g of maize held in 12 cm high x 6.5 cm diameter glass jars covered with ventilated lids. Twenty-three-day old unsexed insect pairs were then introduced into each dish and exposed to treatments. Actellic dust was used as a positive control and all tests were done in three replicates. Insects were considered dead when probed with sharp objects and there were no responses [32]. The number of dead insects in each vial was counted after 21 days after treatment to estimate mortality as follows:
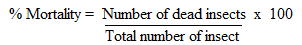 Data on percentage adult weevil mortality were corrected using Abbott’s formula [33]: PT = (Po – Pc) / (100 - Pc) Where PT = Corrected mortality (%); Po = Observed mortality (%); PC = Control mortality (%).
Data on percentage adult weevil mortality were corrected using Abbott’s formula [33]: PT = (Po – Pc) / (100 - Pc) Where PT = Corrected mortality (%); Po = Observed mortality (%); PC = Control mortality (%).2.7. Growth Inhibition Assay
- The test was done according to Ileke and Oni [32] with some modifications. 20 g of clean undamaged and uninfected corn grains were placed in 12 cm high x 6.5 cm diameter glass jars glass jars. Test materials (crude extracts and isolated compounds) were thoroughly mixed with the grains in each jar. Crude extracts and pure compounds were mixed with talc thoroughly before being applied to the grains [34]. A mixture of twenty-seven-day old unsexed maize weevils was introduced in each jar and covered with filter paper [31]. The female adults were allowed to oviposit on the seeds for 4 days. On day 5, all insects were removed from each container and the seeds returned to their respective containers. Progeny emergence (F1) was recorded at six weeks (42 days). The containers were sieved out and newly emerged adult weevils were counted [32]. At week six, the grains were reweighed and the percentage loss in weight was determined as follow:
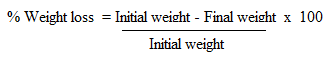 The results from the experiments were expressed as mean ± SD (n=3). Data obtained from the experiments were subjected to analysis of variance and means were separated by least significant difference (LSD) at five percent significant level.
The results from the experiments were expressed as mean ± SD (n=3). Data obtained from the experiments were subjected to analysis of variance and means were separated by least significant difference (LSD) at five percent significant level.3. Results and Discussion
3.1. Compounds Isolated from the Plant
- Chromatographic fractionation of extracts from W. ugandensis afforded seven compounds (Figure 1) namely 7α-acetylugandensolide (1), ugandensolide (2), polygodial (3), warbuganal (4), ugandensidial (5), mukaadial (6) and muzigadial (7). The structures of the compounds were determined using spectroscopic and physical methods. 7α-Acetylugandensolide (1) was isolated as colorless gummy material, melting point 228-230°C, [α]D +24° (MeOH, c, 0.01). IR spectrum showed the presence of ester carbonyl, lactone and olefinic bond at 1732, 1680 and 1632 cm-1 respectively. 13C NMR spectrum exhibited 19 distinct carbon signals corresponding to five methyl (δ 20.6, 20.8, 21.3, 22.9 and 33.1), four methylene (δ 18.3, 36.3, 43.1 and 70.2), three methine (δ 50.5, 66.6 and 69.6) and seven quaternary (δ 33.3, 35.4, 140.3, 150.6, 169.8, 170.9 and 171.1) carbon atoms (Table 1). The 13C NMR peaks at δ 170.9 and 70.2 were attributed to the lactone carbonyl and methylene carbons at C11 and C12 respectively. The 13C NMR peaks at δ 171.1, 169.8, 21.3 and 20.8 were attributed to the two acetoxy groups at C6 and C7. Peaks at δ 69.6 and 66.6 which correlated to protons δ 5.54 (tr, J = 1.7 Hz Hz) and 5.15 (dd, J = 1.3 Hz) in the HMBC spectrum were assigned to the oxymethine carbons at C6 and C7 respectively. The 13C NMR peaks at δ 150.6 and 140.3 were assigned to the double bond carbons at C8 and C9 respectively. The stereochemistry at C-6 and C-7 followed from the small vicinal coupling constants of protons at H-5, H-6 and H-7 [35]. High resolution mass spectrum (70 eV) displayed a molecular ion peak at m/z 350.1154 [M]+ corresponding to C19H26O6 formula. Spectral data of compounds 2-7 (Table 1) were in agreement with those previously reported for the compounds [35,36].
|
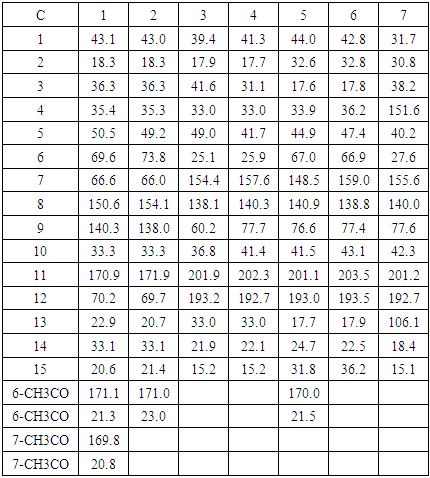
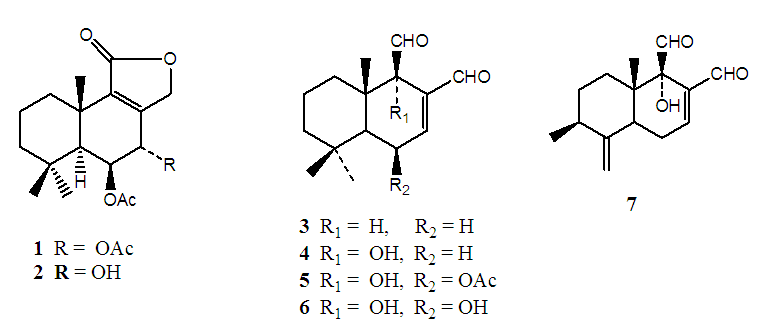 | Figure 1. Compounds isolated from Warburgia ugandensis |
3.2. Repellent Activity
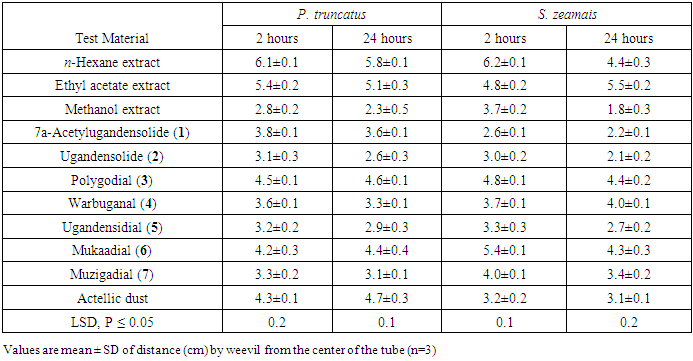
- The repellent activity of crude extracts and compounds from W. ugandensis against the insect pests was recorded after 2 and 24 hours of exposure and the results were as presented in Table 2. All the extracts tested exhibited repellence activity against the test insects. Among the crude extracts, n-hexane extract was the most repellent against both insects followed by ethyl acetate and methanol extracts. Hexane extract had mean repellence ranging from 5.8 to 6.1 cm and 4.4 to 6.2 cm against P. truncatus and S. zeamais respectively. All the compounds isolated also caused some repulsion against the insects. Polygodial (3) was the most repellent compound against P. truncatus followed by mukaadial (6) with mean repellence ranging between 4.5-4.6 and 4.2-4.4 cm respectively. S. zeamais was most repelled by mukaadial (6) and polygodial (3) and the two compounds exhibited mean repellence ranging between 4.3-5.4cm and 4.4-4.8 cm respectively.
|
3.3. Mortality, Growth Inhibition and Weight Loss Prevention Activities
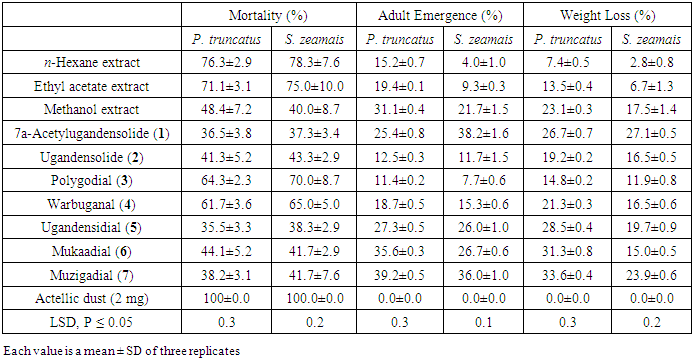
- The crude extracts and compounds were tested for mortality and growth inhibition and grain weight loss prevention activities against P. truncatus and S. zeamais and the results were as presented in Table 3. Both P. truncatus and S. zeamais were more susceptible to hexane and ethyl acetate extracts than the methanol extract. Mortality activity of hexane ranged between 76.3-78.3% while that of ethyl acetate extracts ranged between 71.1-75.0%. Among the compounds isolated from the plant, polygodial (3) and warbuganal (4) were the most toxic to the insects. Mortality activity of polygodial (3) ranged between of 64.3-70.0 while that of warbuganal (4) was 61.7-65.0%.
|
4. Conclusions
- Plant material and extracts have been used to control insect pests. The use of plant materials as pesticides is considered to be environmentally safe. Furthermore, plant materials are readily available, renewable and chances of insects developing resistance are negligible. Future studies aimed at determining synergism and antagonism effects of the bioactive compounds from W. ugandensis are necessary to determine the combinations that offer the best control against insect infestation of stored Zea mays.
ACKNOWLEDGEMENTS
- The authors are grateful to the National Commission for Science, Technology and Innovation (NACOSTI) and Biosciences Eastern and Central Africa Network (BecANet) for financial support.