-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
International Journal of Biophysics
p-ISSN: 2168-4979 e-ISSN: 2168-4987
2016; 6(1): 7-12
doi:10.5923/j.biophysics.20160601.03

Hologram Formation on the Human Eye Retina
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLTitar V. P. , Ielchishcheva Yu. V.
Laboratory of Radio and Optical Holography, V. N. Karazin Kharkiv National University, Kharkiv, Ukraine
Correspondence to: Titar V. P. , Laboratory of Radio and Optical Holography, V. N. Karazin Kharkiv National University, Kharkiv, Ukraine.
| Email: |  |
Copyright © 2016 Scientific & Academic Publishing. All Rights Reserved.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Now, there exists no convincing explanation of illogical structure of higher animal inverted eyes. The existing well-known models of visual perception usually assume that the fields registered by the visual receptors are focused images of objects. In present paper, we propose an improved model of the holographic data formation on the human eye retina with subsequent image reconstruction. The possibility of the hologram formation on the retina is shown. The hologram is formed due to the interference of waves recorded by rods and cones, and waves that have passed through the intrinsically photosensitive retinal ganglion cells (ipRGCs). In the resonant quasi-crystal medium, the nonlinear signal interactions between object signals generated by the rods and cones, and the reference signal generated by the ipRGCs with a time delay between the signals is taken into account. The theoretical analysis of complex holograms formation on the retina is done. The results allow us to better understand the processes of visual perception and they can be used to develop new methods of eye disease diagnostic and treatment, and also can be applied to creation of new information systems: holographic TV and atmospheric laser communication.
Keywords: Human visual system, Retina, Holographic model, Intrinsically photosensitive retinal ganglion cells
Cite this paper: Titar V. P. , Ielchishcheva Yu. V. , Hologram Formation on the Human Eye Retina, International Journal of Biophysics , Vol. 6 No. 1, 2016, pp. 7-12. doi: 10.5923/j.biophysics.20160601.03.
Article Outline
1. Introduction
- Until now, there is no convincing explanation of illogical structure of inverted eyes of higher animals. The light sentient terminal elements of photoreceptor cells are situated at a maximal distance from the lens, and rest against the dark pigment layer. Therefore, the light incident upon the eye first passes through the layers of retinal nerve fibers and then is detected by the photosensitive receptors.In contrast to the well-known models, in which fields registered by visual receptors are interpreted as focused image of the object, we considered the possibility of a hologram formation on the retina of the human visual system and subsequent reconstruction of a three-dimensional image. At first, we have assumed and then have shown that the eye optical system forms not a focused image of an observed object on the retina, but its spatial spectrum. A coding is performed by anisotropic active quasi-crystalline structures of cone iodopsin and rod rhodopsin [1].During recent years, there appeared publication in which analogy between processes in the human brain during the visual information processing and holographic processes can be traced. Most authors believe that quasi-holographic processing of visual information can be carried out in neuron modules of the visual cortex [2], or even at the retina level [3]. In addition, intensive investigations of nonlinear anisotropic bio-crystal structures are now started [4]. Studies of the bacteriorhodopsin, which property is similar to that of the visual rhodopsin, the eye photosensitive pigment, have shown that these materials can be used as a medium for the nonlinear dynamic holography [5]. There exist same psychophysical and sensor physiological features, which cannot be explained in framework of the existing models.In 1986, based on the analysis of hypotheses and models of visual perception, we offered a hypothesis concerning the holographic mechanism of the visual perception [6]. This hypothesis has been theoretically proved and verified by simulations of visual illusions [7]. Thus, we have come to the conclusion that the visual processes are of holographic nature. Then, a question arose of how the hologram can be formed on the retina. We have proposed and verified several models, in particular, a model of an intensity hologram formation [8], which has been improved and suggestion was made that the retinal photoreceptors record a picture of the observed object as a hologram. The field reflected from the sight fixing point, the so-called bright point, that contains phase defined by the distance to the object. This field can be used as a reference signal for the hologram formation on the retina. Defocused fields received from the surrounding area the bright point can used as the object field, since it carries the amplitude-phase information concerning an extended area of the object [9, 10]. The proposed model of the visual perception allows us to understand the processes of visual perception. Based on the model of the visual perception, new methods of eye disease diagnostic and treatment were developed, namely: a method of holographic macular stimulation [11] and a method of holographic visual contrast measurement [12]. Furthermore, we have proposed new principles of information systems [13] development: the holographic television [14, 15] and laser communication systems with an open resonator [16, 17].After the discovery of intrinsically photosensitive retinal ganglion cells (ipRGCs) [18] in the retina of mammals, we propose to improve the model by stating that the reference signal can be formed by the ipRGCs when light passes through them. The present work is aimed at the theoretical justification of the advanced model of the visual perception.
2. The Human Eye Retina
- The retina is the light-sensitive inner membrane of the eye. It has a complicated multilayer structure [19, 20]. The internal layer, closest to the pupil, is the layer of nerve fibers (fibers of the visual nerve). Then the ganglion cell layer, inner synaptic layer, bipolar cell layer, and outer synaptic layer follow one after the other. The layer of receptor cells, which is adjacent to the pigment layer, which is the most distant from the pupil, is composed of two types photoreceptor with different functionality: rod and cone. There are from 110 to 123 million rods and from 6 to 7 million cones in the human eye retina. Distributed irregularly over the retina. A yellow spot (macula) of an oval form with axes 2 mm and 0.8 mm can be found in the center of the retina [21]. In the middle of the macula there is a foveal region, or fovea centralist filled with cones only. In the fovea connections between each cone and bipolar or ganglion cells are more direct than at the periphery.The regions without rods occupy from 1.70 to 9.5 degree of the visual space. In the center of the retina the region, which diameter is one degree, the visual function is ensured entirely by cones. The rod vision can be neglected within two central degrees [22].There are about 25,000 cones in the central rod-free foveola [23]. The size and shape of the receptors are varying in a wide range at various retina regions. For example, the cone diameter is 2 µm in the foveal area, while at the periphery it may reach up to 6 µm.The light propagates through the system of nerve fibers located over cones in a plane parallel to the macular. The nerve fiber layer due to its structure becomes form birefringent [24]. The light transmitted through the medium decomposes into two linearly polarized waves, ordinary and extraordinary, with mutually orthogonal linear polarizations. In the human eye retina, an effective method of reducing video data redundancy is realized, since the number of hologram elements is reduced due to the non-equidistant arrangement of the cones in the macula. The cone photoreceptors filtrate the incident light and discriminate three quasi-monochromatic components: red, green, and blue, according to the type of iodopsin pigment in the cones [25].
3. Formation of the Reference Wave by the ipRGCs
- In 2002, Berson et al. [18] discovered a third type of photoreceptor cell in the ganglion cell layer of the mammalian retina. The axons of these cell form a nerve fiber layer. This new type of photoreceptor cells, intrinsically photosensitive retinal ganglion cells (ipRGCs) are classified into five subtypes [26]. The sensitivity of the ipRGC photoreceptor depends on the wavelength of the light and, hence, on the light color [27]. The ipRGCs can also regulate many biological functions, such as the circadian rhythms, body temperature, cardiac rhythm, cortisol and melatonin synthesis [28-30]. Since the cone has circular shape, a pointwise description of the retinal image is carried out, depicted by very fine mosaics, consisting of excited neurons. At the same time the dendrites of the imRGCs are spread more widely in the plane of the retina than in the outer segments of the classical photoreceptor, ensuring, a greater spatial integration of the signals. Melanopsin-positive ganglion cells can be found throughout the retina, though a slight density increase is observed in the retina center [31]. The dendrites of these cells form plexuses, intensively overlapping in the inner plexiform layer [32, 33]. Dendritic arbors of the separate melanopsin-positive ganglion cells are rather wide, covering about 500 μm or 15° [18], while the rod and cone outer segments cover only about 1 μm or 0,05°.
4. Hologram Formation
- The light sensitive imRGCs combine the direct responses to light stimuli based on melanopsin, photopigment in these cells [34, 35], with the signals received from the rods and cones, and transfer them directly to a subset of retinal targets in the brain [26, 36].A new holographic model of visual perception is proposed on the basis of these properties of ganglion cells.Since the retina layers have a clearly expressed anisotropy, the layer of nerve fibers acts as a positive uniaxial crystal, and the outer segments of the rods and cones behave as negative uniaxial crystals. The optical axes of these crystals are mutually perpendicular. When an arbitrary polarized light propagates through such optical system, a set of four linearly polarized waves: two ordinary and two extraordinary waves are registered by the cones in the outer segments of the retina photoreceptors for the R, G, and B components of quasi-monochromatic white light.Registration of complex Fourier hologram on the retina is performed by two reference waves with mutually orthogonal polarizations, which are formed by ipRGCs and selected by the retina layers from the waves that passed through the same layer of the nerve fibers and are shifted in phase relative to one another at
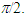 These signals can interfere if they are monochromatic, have the same polarization and are registered simultaneous. The rod and cone signals are delayed for several microseconds, since a cascade of biochemical reactions take place in the photoreceptors, and the electrical signal generated in response to photon absorption can be delayed [37].It should be noted that molecules of the visual pigments packed in discs can be thought as a dynamic recording medium [38, 39], since the light stimulus causes an instantaneous rearrangement of protein globules of chromoproteids that causes a rapid change of the refractive index. The variation of the refractive index caused by any part of the photon flow passing through the dynamic medium influences the propagation conditions for the other photons. In other words, processes of images writing and reading, which are related in time and are characteristic of the dynamic holography, occur in the outer segments of photoreceptor cells. Any portion of light beam that hit the retina is limited by arbitrarily selected time interval and varies due to the refractive index variation caused by the previous part of this beam. That is, the light beam reads the image recorded recently and writes a new image, that corresponds to the intensity distribution of the light beam, arising at the time of recording, and so on. In addition, the retina is a resonant quasi-crystalline medium, wherein the absorption-emission line produced by transition to the lower energy level coincides with the wavelength of the incident light [40]. It is known that a photon echo can be observed in such a medium. The echo can be interpreted as the ability to store and render temporal parameters of the electromagnetic field, such as the time interval between successive light signals. The medium can also remember the spatial phase distributions of the first signal.On the basis of the photon echo effect, Shtyrkov [41], have shown that the dynamic hologram can be registered in the resonant medium, even if the object and reference waves do not coincide in time. Let the object wave incident on the resonant medium
These signals can interfere if they are monochromatic, have the same polarization and are registered simultaneous. The rod and cone signals are delayed for several microseconds, since a cascade of biochemical reactions take place in the photoreceptors, and the electrical signal generated in response to photon absorption can be delayed [37].It should be noted that molecules of the visual pigments packed in discs can be thought as a dynamic recording medium [38, 39], since the light stimulus causes an instantaneous rearrangement of protein globules of chromoproteids that causes a rapid change of the refractive index. The variation of the refractive index caused by any part of the photon flow passing through the dynamic medium influences the propagation conditions for the other photons. In other words, processes of images writing and reading, which are related in time and are characteristic of the dynamic holography, occur in the outer segments of photoreceptor cells. Any portion of light beam that hit the retina is limited by arbitrarily selected time interval and varies due to the refractive index variation caused by the previous part of this beam. That is, the light beam reads the image recorded recently and writes a new image, that corresponds to the intensity distribution of the light beam, arising at the time of recording, and so on. In addition, the retina is a resonant quasi-crystalline medium, wherein the absorption-emission line produced by transition to the lower energy level coincides with the wavelength of the incident light [40]. It is known that a photon echo can be observed in such a medium. The echo can be interpreted as the ability to store and render temporal parameters of the electromagnetic field, such as the time interval between successive light signals. The medium can also remember the spatial phase distributions of the first signal.On the basis of the photon echo effect, Shtyrkov [41], have shown that the dynamic hologram can be registered in the resonant medium, even if the object and reference waves do not coincide in time. Let the object wave incident on the resonant medium 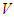 at time
at time 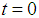 and transfers part of the atoms from the lower ground state 1 to the upper, excited state 2. The photon echo theory predicts that in this state, the phase of the atom oscillation within the transverse relaxation time is correlated with the phase of the object wave during its interaction with the medium. Thus, the nonlinear interaction of the object signals formed by the cones and rods and the reference signals of the ipRGCs), a hologram can be recorded in the resonant medium even if a time delay between these signals exists. The wavefront of the reference waves across the retina surface can be maintained constant by the lens accommodation muscles [1] and the eye muscles [42]. Each of the quadrature hologram components is an interference structure, formed by the interaction of the object
and transfers part of the atoms from the lower ground state 1 to the upper, excited state 2. The photon echo theory predicts that in this state, the phase of the atom oscillation within the transverse relaxation time is correlated with the phase of the object wave during its interaction with the medium. Thus, the nonlinear interaction of the object signals formed by the cones and rods and the reference signals of the ipRGCs), a hologram can be recorded in the resonant medium even if a time delay between these signals exists. The wavefront of the reference waves across the retina surface can be maintained constant by the lens accommodation muscles [1] and the eye muscles [42]. Each of the quadrature hologram components is an interference structure, formed by the interaction of the object 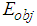 and
and 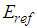 reference waves can be written as
reference waves can be written as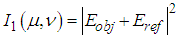 | (1) |
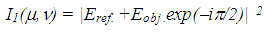 | (2) |
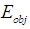 is defined by the relation
is defined by the relation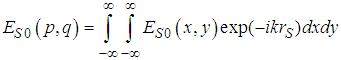 | (3) |
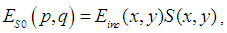
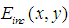 is the complex amplitude of the light incident on the object, and
is the complex amplitude of the light incident on the object, and 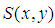 is the object reflection function.After some mathematics [43], we can write the expressions (1) - (2) as
is the object reflection function.After some mathematics [43], we can write the expressions (1) - (2) as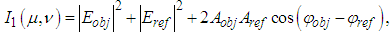 | (4) |
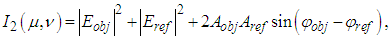 | (5) |
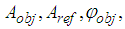 and
and 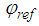 are the amplitudes and phases of the object and reference waves. After averaging the signal over the retina and subtracting the mean from each signal registered by cone photoreceptor, the interference component of the hologram signal will be recorded by the cone photoreceptors. Therefore, we consider only the sine and cosine terms in (4) - (5) and the model of the complex hologram can be presented as
are the amplitudes and phases of the object and reference waves. After averaging the signal over the retina and subtracting the mean from each signal registered by cone photoreceptor, the interference component of the hologram signal will be recorded by the cone photoreceptors. Therefore, we consider only the sine and cosine terms in (4) - (5) and the model of the complex hologram can be presented as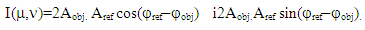 | (6) |
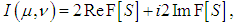 | (7) |
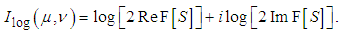 | (8) |
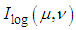 overlaid by a spatial-frequency filter
overlaid by a spatial-frequency filter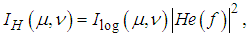 | (9) |
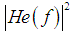 is the spatial-frequency characteristic of the eye [47], which is the transfer function, formed by a set of the channels (filters) of the visual cortex, tuned to a specific spatial frequency [48].The image reconstruction can be done by the inverse Fourier transform which is carried out in the visual cortex. The restored image can be written as
is the spatial-frequency characteristic of the eye [47], which is the transfer function, formed by a set of the channels (filters) of the visual cortex, tuned to a specific spatial frequency [48].The image reconstruction can be done by the inverse Fourier transform which is carried out in the visual cortex. The restored image can be written as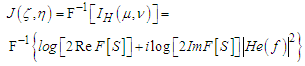 | (10) |
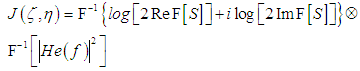 | (11) |
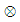 is the convolution operator and
is the convolution operator and 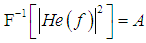 is the instrumental function of the optic tract. Thus, the reconstructed image (11) is the convolution of the inverse Fourier transform of the complex hologram and the instrumental function of the optic tract
is the instrumental function of the optic tract. Thus, the reconstructed image (11) is the convolution of the inverse Fourier transform of the complex hologram and the instrumental function of the optic tract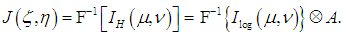 | (12) |
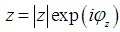 | (13) |
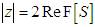 and
and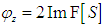 . Let this function be related to a complex function
. Let this function be related to a complex function 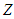 by the Fourier transform
by the Fourier transform 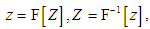 then the classical complex cepstrum analysis can be applied to our model, and the reconstructed image can be presented as a convolution of the complex cepstrum of the function Z and the instrumental function of the optic tract
then the classical complex cepstrum analysis can be applied to our model, and the reconstructed image can be presented as a convolution of the complex cepstrum of the function Z and the instrumental function of the optic tract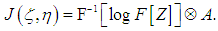 | (14) |
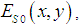 then the object image will be restored. Subsequent processing of visual information can be made both in the higher subdivisions of the visual cortex and during the reverse signal transmission from the cortex to the eye retina with the image imposition to the outer space in accordance with the Sechenov’s principle of relatedness [49], which may also be explained based on the holographic model of the visual system. We may suppose that dynamic hologram [39] is formed in the retinal layers, which ensures the wave front reversal, and, hence, overlapping of the reconstructed image with the shape of the observed object.The second stage of the spatial information processing is the stage of nervous activity, which begins on the retina. The incident electromagnetic wave is detected by the mosaic of photoreceptor and the signals through the neural network are transmitted to the ganglion cells and nerve fibers and to the neurons of the visual segment of the cerebral cortex. The nature of this processing stage in the nervous system is known only partially.Thus, in accordance with the model, when the signals from the visual cortex of the brain fall into the dynamic hologram, the wave front reversal, the image imposition of the observed object in outer space and superimposition of the image with the object shape can occur.
then the object image will be restored. Subsequent processing of visual information can be made both in the higher subdivisions of the visual cortex and during the reverse signal transmission from the cortex to the eye retina with the image imposition to the outer space in accordance with the Sechenov’s principle of relatedness [49], which may also be explained based on the holographic model of the visual system. We may suppose that dynamic hologram [39] is formed in the retinal layers, which ensures the wave front reversal, and, hence, overlapping of the reconstructed image with the shape of the observed object.The second stage of the spatial information processing is the stage of nervous activity, which begins on the retina. The incident electromagnetic wave is detected by the mosaic of photoreceptor and the signals through the neural network are transmitted to the ganglion cells and nerve fibers and to the neurons of the visual segment of the cerebral cortex. The nature of this processing stage in the nervous system is known only partially.Thus, in accordance with the model, when the signals from the visual cortex of the brain fall into the dynamic hologram, the wave front reversal, the image imposition of the observed object in outer space and superimposition of the image with the object shape can occur.5. Conclusions
- 1. An improved holographic model of visual perception is proposed, which is based on an assumption that the two quadrature components of the spatial spectrum are formed by the visual receptors. The model includes the signal averaging over the retina, subtraction of the averaged signal, logarithmic operations, spatial filtering, and the signal saturation. Description of the logarithmic processing of the spatial spectrum quadrature components in the visual system and the image reconstruction is done by using the complex cepstrum analysis. 2. The image obtained after the log processing and bandpass spatial filtering is the convolution of the complex Fourier cepstrum of the light intensity on the object with the instrumental function of the optic tract. The object image thus obtained can be presented as a certain visual stimulus consisting of the object contours and vertexes, which can be used by the visual system, for image identification and for determining its shape, dimensions and position in space.3. If the visual system can perform the inversion of the complex cepstrum, the field forming the object image, i.e., the brightness distribution on the object, can be imposed into object space using the wave front reversal technique.4. The proposed model brings us to an understanding of the visual perception processes and can be used both in systems for eye diseases diagnostic and in systems for registration and processing of electromagnetic fields, in particular, for holographic television and laser communication systems.