-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
Advances in Life Sciences
p-ISSN: 2163-1387 e-ISSN: 2163-1395
2015; 5(3): 53-57
doi:10.5923/j.als.20150503.01
Studies of in vitro Adventitious Root Induction and Flavonoid Profiles in Rumex crispus
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLMajid Mahdieh1, Mitra Noori1, Simin Hoseinkhani2
1Department of Biology, Faculty of Science, Arak University, Arak, Iran
2MSc student of Department of Biology, Faculty of Science, Arak University, Arak, Iran
Correspondence to: Mitra Noori, Department of Biology, Faculty of Science, Arak University, Arak, Iran.
| Email: |  |
Copyright © 2015 Scientific & Academic Publishing. All Rights Reserved.
Flavonoids are the principal bioactive components of the medicinal plant Rumex crispus, to which various diverse pharmacological properties are attributed. Adventitious root culture of leaf explants of R. crispus was established using different MS media supplemented by different concentrations of Auxins and a combination of Naphthalene Acetic Acid (NAA) and Kinetin (Kn) for growth and flavonoids production. NAA was more effective than Indole Acetic Acid (IAA) to induce adventitious roots. 5 µM NAA treatments had the most number of roots. Maximum root length and root fresh weight were obtained in 0.5 µM Kn. 0.5 µM Kn using in combination with NAA in culture medium increased root fresh weight. Results of 2-dimensional paper and thin layer chromatography of the samples extracts showed that hormons are influenced flavonoids in culture medium. The combination of NAA and Kinetin in culture medium reduced or suppressed Myricetin and Naringenin production and increased number of aglycones. Quercetin was not found in root on medium containing 0.5 µM Kn. Isorhamnetin, Kaempferol, Rhamnetin and Rutin were found in all of treatments that had produced roots. It is believed that root flavonoids are related to root growth, development and differentiation in R. Crispus species.
Keywords: Adventitious root, Auxin, Cytokinin, Flavonoids, Rumex crispus
Cite this paper: Majid Mahdieh, Mitra Noori, Simin Hoseinkhani, Studies of in vitro Adventitious Root Induction and Flavonoid Profiles in Rumex crispus, Advances in Life Sciences, Vol. 5 No. 3, 2015, pp. 53-57. doi: 10.5923/j.als.20150503.01.
Article Outline
1. Introduction
- Rumex crispus is an herbaceous perennial weed plant, belongs to Polygonaceae which has been distributed widely in the humid regions of northern hemisphere including Iran, mostly in the acidic (silicate) soils. Traditionally, its roots have been largely recommended by herbalists for range of skin diseases [1]. The plant contains many bioactive substances such as flavonoids, anthraquinones (particularly in roots), tannins, saponins and triterepenoids [2-4]. Flavonoids possess a wide range of biological activities, medicinal and pharmacological effects [5].Plant cell culture technologies developed in the past as possible tools for secondary metabolites production. In most cases, for the large scale production of these compounds, production is too low for commercialization. Secondary metabolites production is under strict metabolic regulation and tissue specific localization. Hence the differentiated cultures such as root cultures are widely studied. Roots of numerous plant families are the site for biosynthesis or accumulation of major secondary metabolites. In vitro root culture has become an alternative method for the production of valuable secondary metabolites on commercial scale [6]. Transformed root (hairy root) cultures produced by Agrobacterium rhizogenesis offer a promising alternative method to cell culture, but hairy roots usually produce opine-like substances which are lethal to mammalian cells [7] and also GMOs are not always accepted in several countries like Australia, Japan, France, Korea, New Zealand etc. Adventitious roots induced by in vitro methods showed high rate of proliferation and active secondary metabolism [8, 9]. Adventitious roots are natural, grown vigorously in phytohormone supplemented medium and have shown tremendous potentialities of accumulation of valuable secondary metabolites [10]. It was reported that adventitious root induction technique has been applied on economically important plants [11, 12]. For example, mass production of isoflavonoids through adventitious root cultures of Iris germanica had been conducted by Tomoyoshi et al (2005) who noticed that its adventitious root cultures accumulated a high concentration of isoflavones that capable to be utilized in medicinal production [12]. However, optimization and scale-up is required to increase root biomass and secondary metabolites [13-16].Therefore, the purpose of the study was to develop an efficient in vitro development of root cultures from leaf explants of Rumex crispus, an important medicinal plant. In the present work, we have established a reproducible method for high frequency root induction, root growth from leaf explants on MS solid medium and investigated the effects of Auxin and Cytokinin on flavonoid constitutes of in vitro adventitious roots.
2. Materials and Methods
2.1. Plant Material and Adventitious Root Culture Establishment
- In vitro cultured plantlets of Rumex crispus were established from seeds (collected from Arak, Iran, 34° 06′ N, 49° 46′ E during 2013) on basal Murashige and Skoog (MS) medium [17] and were used as plant material in this study. Leaves which were excised from in vitro grown plantlets at the age of 8 weeks, cut into small segments (0.7 cm×0.7 cm) and cultured on MS medium (3% sucrose) supplemented with different concentrations (0, 2.5, 5 and 10 µM) of Indole-3-Acetic Acid (IAA) or 1-Naphthalene Acetic Acid (NAA). The second experiment involved combining 2.5 or 5 µM NAA with different concentrations of Kinetin (0, 0.1, 0.5 µM). All the cultures were maintained for 3 weeks at 25 ± 1°C in dark. The rooting percentage (%), fresh weight (mg), root length (cm) and the number of roots formed per explant were recorded after 3 weeks.
2.2. Preparation of Root Extract
- A ground-dried root (0.2 g) was refluxed with 10 ml of 80% methanol (MeOH) at 80 °C for 1 h. The mixture was cooled and after centrifugation at 4000g for 10 min, the supernatant solution was filtered. The extract evaporated to dryness by rotary evaporation at 40°C and taken up in 2 ml of 80% MeOH for analysis by 2-dimensional paper chromatography (2D PC) and thin layer chromatography (TLC) based on available references [18, 19].
2.3. Isolation and Identification of Flavonoids by 2-DPC
- For the detection of flavonoids, ca 20 µl of each extracts was applied to chromatography paper as a concentrated spot (10 applications of 20 µl). The chromatogram for each sample was developed in BAW (n-BuOH-HOAc-H2O=4:1:5; v/v) for first direction and in HOAc (=15% aqueous acetic acid) for second direction. After development, the chromatograms were viewed in long wave UV light (366 nm) and fluorescent spots were marked. Rf values in BAW and 15% HOAc were calculated.
2.4. Identification of Flavonoids Types
- When sufficient amounts of purified flavonoids had been obtained, they were identified by means of UV spectrophotomerty using shift reagents to investigate the substitution patterns of the flavonoids [18, 19] and by acid hydrolysis to identify the aglycone and sugar moieties. The TLC plates were run in three solvents alongside standards to identify the aglycone moiety [20, 21]. Co-chromatography with standards was also performed where possible. Flavonoid standards available for comparison during the study obtained commercially from Merck, Sigma and Fluka).
2.5. Statistical Analysis
- Data were analyzed by One-way analysis of variance (ANOVA) to detect significant differences between means. Means differing significantly were compared using the Duncan’s multiple range tests at the 5% probability level.
3. Results and Discussion
3.1. Root Formation from Different Explants
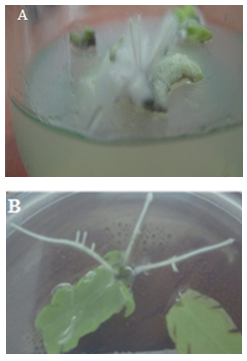
- We tested various explants including root, stem, petiole and leaf segments to find the best explant to adventitious root initiation. Adventitious root cultures were not established by the root (Figure 1a) and stem explants, because of slow growth and development of adventitious roots from both the explants. Thus, the leaf and petiole explants were alternatively used to establish the root culture of curled dock plant. Adventitious roots were fully developed from the leaf explants within 2 weeks (Figure 1b) followed by petiole explants (3 weeks) (Figure 1c). So leaf explants was found to be the best explants in root induction.
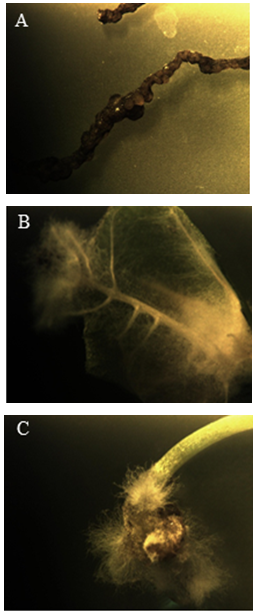 | Figure 1. The effects of NAA (5µM) on the formation of adventitious roots on the segments of Rumex crispus within 2-3 weeks. A. Less roots occurred at the main root, B. Highly branched roots formation on leaf explants, C. Thin and short roots in petioles |
3.2. Effect of Exogenous Auxins and Cytokinin on Root Production
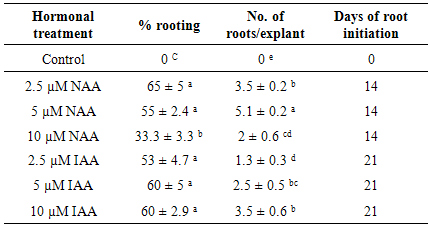
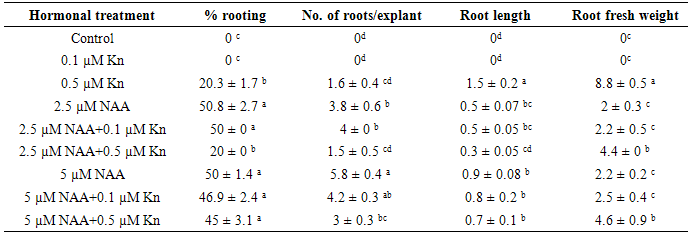
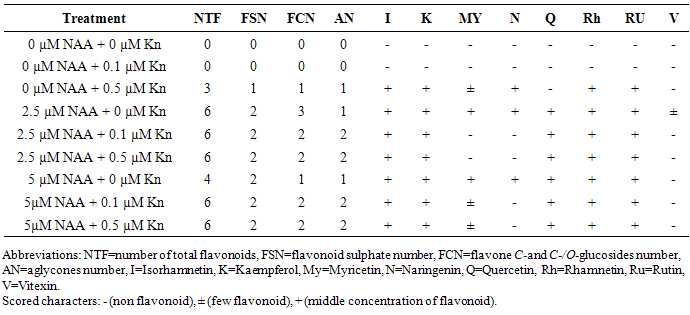
- Auxins promote the formation of adventitious roots on explants. Among the NAA concentrations, supplementation of 2.5 µM NAA to culture medium was generated the highest rooting percentage (65%) within 2 weeks, which was followed by 5 µM NAA (55%). Contrarily, root initiation was suppressed in 10 µM NAA (Table 1). The highest root number per explant was achieved in 5 µM NAA within 2 weeks. IAA also found to be successfully induced the formation of adventitious roots, and the highest rooting ability for IAA was achieved in 10 µM within 3 weeks (Table 1). Contrary to our result, in Malus [22] IAA was effective for the production of roots and NAA strongly inhibited the growth of roots. This is in line with the fact that a particular type of Auxin is effective in enhancing rooting in particular species [23]. Thus we selected 2.5 and 5 µM concentrations of NAA for further experiments on effects of Cytokinin in combination with NAA on rooting and root biomass production in explants. The highest number of roots and rooting percentage were obtained in leaf explants cultured on MS media with 5 µM NAA, but by the reason very low root weight, this medium was not suitable for rooting (Table 2, Figure 2a). The influence of NAA alone on root growth (fresh weight) was negligible. The addition of Kinetin (0.5 µM) significantly increase root weight in treatments containing NAA, which means Kinetin could promote the growth of the roots, however the root number was decreased in MS media containing 0.5 µM Kn.
|
 | Figure 2. Adventitious root formation from in vitro leaf explants in the medium containing: A. 5 µM NAA or B. 0.5 µM Kn |
|
3.3. Effect of Exogenous Auxin and Cytokinin on Flavonoids Production
- The type and concentration of Auxin and Cytokinin, either alone or in combination, has been known to strongly influence growth as well as the secondary metabolites in tissue culture. Flavonoids were isolated and identified by 2-dimensional paper and thin layer chromatography. Results of total flavonoids (Table 3) showed that maximum number of total flavonoids (=6) was found in roots on media containing 2.5 µM NAA alone or in combination of Kinetin followed by roots obtained on 5 µM NAA. Addition of 0.5 µM Kn showed a beneficial effect for biomass, as well as flavonoid production. Addition of Auxin (2, 4-D) and Kn into the media were also found to enhance the flavonoids production in Genista tinctoria [27]. According to Maurmann et al (2006), the presence of Kn in combination with 2, 4-D was beneficial to valepotriates accumulation in Valeriana gelechomifolia callus culture [28]. Roots obtained on 0.5 µM Kn were found to contain the least amount of total flavonoids (=3). The flavonoid composition also was influenced by hormone combinations in culture media. The combination of NAA and Kinetin in culture medium reduced or suppressed Myricetin (My) and Naringenin (N) production and increased number of aglycones. Quercetin was not found in root on medium containing 0.5 µM Kn. Isorhamnetin, Kaempferol, Rhamnetin and Rutin were found in all of treatments that had produced roots. There was not Vitexin in all of roots with the exception of 2.5 μM NAA treatment (Table 3). All of studied samples lacked Apigenin, Chrysin, Genistein, Luteolin, Morin and Tricin.As Curir et al (1990) showed in reproduced Eucalyptus gunnii Hook plants from lateral bud stimulation in vitro method, flavonoids association and adventitious root production have direct relationship together and both are overshadowed hormonal treatment [29]. It is believed that root flavonoids are related to root growth, development and differentiation in this species.
4. Conclusions
- This study has revealed the potential of in vitro adventitious roots as a source for flavonoids production in this species. The type and concentration of Auxin and Cytokinin, either alone or in combination strongly influence growth as well as the secondary metabolites in tissue culture. For example NAA was more effective than IAA to induce adventitious roots in R. crispus. It is believed that root flavonoids are related to root growth, development and differentiation in this species.
ACKNOWLEDGEMENTS
- This research has been financially supported by the Higher Education Commission (HEC) of Arak University, Iran.
References
| [1] | EL-BAKRY AA, MOSTAFA HAM, ALAM AE. 2012. Antioxidant activity of Rumex vesicarius L. at the vegetative stage of growth. Asian J Pharm Clin Res, 5:111-117. |
| [2] | NOORI M, CHEHREGHANI A, KAVEH M. 2009. Flavonoids of 17 species of Euphorbia, (Euphorbiaceae) in Iran. Toxicol Environ Chem, 91: 631-641. |
| [3] | RAO KNV, SUNITHA CH, BANGI D, SANDHYA S, MAHESH V. 2011. A study on the nutraceuticals from the genus Rumex. Hygeia J D Med, 3:76-88. |
| [4] | ORBAN-GYAPAI O, RAGHAVAN A, VASAS A, FORGO P, HOHMANN J, SHAH ZA. 2014. Flavonoids isolated from Rumex aquaticus exhibit neuroprotective and neurorestorative properties by enhancing neurite outgrowth and synaptophysin. CNS NeurolDisord Drug Targets, 13:1458-1464. |
| [5] | NOORI M. 2012. Phytochemicals, A Global Perspective of Their Role in Nutrition and Health. In Tech; Chapter 7, Flavonoids in some Iranian Angiosperms; p. 151-166. |
| [6] | CHANDRA S, CHANDRA R. 2011. Engineering secondary metabolite production in hairy roots. Phytochem Rev, 10:371-395. |
| [7] | YOSHIKAWA T, FURUYA T. 1987. Saponin production by cultures of Panax ginseng transformed with Agrobacterium rhizogenes. Plant Cell Rep, 6:449-453. |
| [8] | HAHN EJ, KIM YS, YU KW, JEONG CS, PAEK KY. 2003. Adventitious root cultures of Panax ginseng C. A. Meyer and ginsenoside production through large scale bioreactor systems. J Plant Biotech, 5:1-6. |
| [9] | YU KW, MURTHY HN, JEONG CS, HAHN EJ, PAEK KY. 2005. Organic germanium stimulates the growth of ginseng adventitious roots and ginsenoside production. Process Biochem, 40: 2959-2961. |
| [10] | MURTHY HN, HAHN EJ, PAEK KY. 2008. Adventitious roots and secondary metabolism. Chin J Biotech, 24: 711-716. |
| [11] | ROWINSY EK, CAZENAVE LA, DONEHOWER RC. 1990. Taxol: a novel investigational anti microtubule agent: Review. J. National Cancer Institute, 82: 1247-1259. |
| [12] | TOMOYOSHI A, MASAYUKI I, TOSHIO A, SHIN-ICHI A. 2005. Isoflavonoid production by adventitious root cultures of Iris germanica (Iridaceae). Plant Biotech, 22: 207-215. |
| [13] | CHOI SM, SON SH, YUN SR, PAEK KY. 2000. Pilot-scale culture of adventitious roots of ginseng in a bioreactor system. Plant Cell Tissue Organ Cult, 62: 187-193. |
| [14] | YU KW, GAO WY, HAHN EJ, PAEK KY. 2001. Effects of macro elements and nitrogen source on adventitious root growth and ginsenoside production in ginseng (Panax ginseng C.A. Meyer). J Plant Biol, 44: 179-184. |
| [15] | KIM YS, HAHN EJ, MURTHY HN, PAEK KY. 2004. Effect of polyploidy induction on biomass and ginsenoside accumulations in adventitious roots of ginseng. J Plant Biol, 147: 356-360. |
| [16] | LEE EJ, MOBIN ME, HAHN EJ, PAEK KY. 2006. Effects of sucrose, inoculums density, auxins and aerator volume on cell growth of Gymmema sylvestre. J. Plant Biol, 49: 427-431. |
| [17] | MURASHIGE T, SKOOG F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant, 15: 473-497. |
| [18] | MABRY TJ, MARKHAM KR, THOMAS MB. 1970. The systematic identification of flavonoids. Springer Verlag, Berlin. |
| [19] | MARKHAM KR. 1982. Techniques of Flavonoid Identification. Academic Press, London. |
| [20] | HARBORNE JB. 1998. Phytochemistry methods. Chapman and Hall, London. |
| [21] | NARAYAN MS, THIMMARAJU R, BHAGYALAKSHMI N. 2005. Interplay of growth regulators during solid state and liquid-state batch cultivation of anthocyanin producing cell line of Daucus carota. Process Biochem, 40: 351-358. |
| [22] | KLERK GJD, BRUGGE JT, MARINOVA S. 1997. Effectiveness of Indole-3-acetic acid, Indole-3-butyric acid, α-Naphthalene acetic acid during adventitious root formation in vitro in Malus. Plant Cell Tissue Organ Cult, 49: 39-44. |
| [23] | PURI S, SHAMET GS. 1988. Rooting of some social forestry species. International Tree Crops J, 5: 63-70. |
| [24] | LEE EJ. 2009. Biomass and bioactive compounds production through bioreactor cultures of adventitious roots in Eleutherococcus koreanum. Ph.D. Thesis, Chungbuk National University, Cheong-Ju, Korea. |
| [25] | HILL RA, TUSKAN GA, BOE AA. 1989. In vitro propagation of Hosta sieboldiana using excised ovaries from immature florets. Plant Cell Tissue Organ Cult, 17: 71-75. |
| [26] | PANICHAYUPAKARANANT P, TEWTRAKUL S. 2002. Plumbagin production by root cultures of Plumbago rosea. J Biotech, 5: 228-232. |
| [27] | LUCZKIEWICS M, GLOD D. 2003. Callus culture of Genista plants– in vitro material producing high amount of isoflavones phytoestrogenic activity. Plant Sci, 165: 1101-1108. |
| [28] | MAURMANN N, DECARVALHO CMB, SILVA AL, FETT-NETO AG, VONPOSER GL, RECH SB. 2006. Valepotriates accumulation in callus, suspension cells and untransformed root culture of Valeriana gelechomifolia. In vitro Cell Dev Pl, 42: 50-53. |
| [29] | CURIR P, VANSUMERE CF, TERMINI A, BARTHE P, MARCHESIN A, DOLCI M. 1990. Flavonoid accumulation is correlated with adventitious roots formation in Eucalyptus gunnii Hook micropropagated through axillary bud stimulation. Plant Physiology, 92: 1148–1153. |