Khoshimov B. L.1, Kuliev O. A.2, Kuylanov B. B.3
1Head of the Department of Clinical Sciences, Doctor of Medical Sciences, Alfraganus University, Uzbekistan
2Dean of the Faculty of Medicine, Doctor of Medical Sciences, Alfraganus University, Uzbekistan
3Assistant Professor, Department of Fundamental Sciences, Alfraganus University, Uzbekistan
Copyright © 2025 The Author(s). Published by Scientific & Academic Publishing.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Abstract
This article studies the effects of chronic stress on hippocampal morphology. The study compared the hippocampal structure and developmental dynamics of laboratory rat pups born to stressed mothers with those of the control group. The results show that in offspring born under stress, a decrease in hippocampal neurocytes, vacuolization of the cytoplasm, changes in the size of the nucleus and cytoplasm, as well as lysis of neurocytes were observed. It was also found that the body weight of stressed rat pups was lower than that of the control group. The data obtained indicate that stress can negatively affect the structure of the hippocampus.
Keywords:
Stress, Hippocampus, Neurocytes, Morphology, Neurogenesis, Pathological changes, Laboratory rats
Cite this paper: Khoshimov B. L., Kuliev O. A., Kuylanov B. B., The Effect of Chronic Stress on Adult Hippocamp Morphology, American Journal of Medicine and Medical Sciences, Vol. 15 No. 12, 2025, pp. 4389-4393. doi: 10.5923/j.ajmms.20251512.43.
Article Outline
In today’s modern society, stress is increasingly becoming one of the most important factors affecting human health. The World Health Organization (WHO) has recognized stress as one of the most pressing psychosomatic problems of the 21st century. Its long-term or persistent effects can cause serious damage not only to mental health, but also to the morphological and functional state of the central nervous system, in particular the hippocampus [2,3,10]. The hippocampus is one of the main structures in the brain, which plays a crucial role in memory formation, learning, emotional regulation, and stress responses.Clinical and experimental studies conducted in recent years have shown that chronic stress leads to neuronal degeneration, reduced synaptic plasticity, and decreased neurogenesis in the hippocampus. This, in turn, leads to memory loss, learning disabilities, impaired concentration, emotional instability, and increased susceptibility to mental disorders [1,9]. In particular, the effect of stress on the hippocampus is considered an important pathogenetic link in the development of age-related cognitive decline, Alzheimer's disease, and other dementias [5,7].In addition, a decrease in hippocampal volume is closely associated with mental disorders such as depression, generalized anxiety disorder, and posttraumatic stress disorder (PTSD). Numerous neuroimaging studies have shown that the average volume of the hippocampus in depressed patients is significantly smaller than in healthy individuals. Such morphological changes limit the brain's ability to adapt (neuroplasticity) and impair its ability to assimilate new information [4,6].The sensitivity of the hippocampus to stress has made it an important object for basic scientific research. Today, determining the morphological effects of stress on the hippocampus, studying its mechanisms, and developing ways to prevent or reduce these changes are priorities in the fields of neurobiology, psychiatry, and morphology.Therefore, this topic is extremely relevant not only from a scientific, but also from a practical and social point of view. Because stress-related diseases are among the main causes of global health problems. The morphological response of the hippocampus to stress directly affects the quality of human life, labor productivity and social adaptability. Modern science suggests the use of meditation, physical exercise, psychotherapy, pharmacological drugs, as well as neuroprotective agents to reduce stress and support hippocampal activity. However, the extent to which the effectiveness of these methods is related to the specific morphological changes occurring in the hippocampus has not yet been fully studied.Therefore, an in-depth study of the morphological changes of the hippocampus that occur under stress, identifying the psychophysiological consequences of these changes and applying them to medical prevention and treatment strategies is an important scientific and practical task for modern medicine.Taking the above into account, we set ourselves the goal of studying the structural and functional state of the hippocampal neural tissue of offspring born to mothers under chronic stress.Materials and methods of study. To achieve the goal, 120 white laboratory rats were used. White laboratory rats were divided into 2 groups. Group 1 consisted of 20 healthy rats as a control group. Mother rats in the control group were given 1.0 ml of saline solution intragastrically every morning. A subclavian catheter was used as a probe. The rats were anesthetized under ether anesthesia on days 7, 14, 21, and 30 after birth. Group 2 was the experimental group, and 20 female white laboratory rats were kept in specially prepared cages to induce experimental stress in them. The rats were exposed to various stress factors (noise and excessive light) for several weeks. This stress model was continued after the rats became pregnant and gave birth to their offspring. To confirm the stress state in animals, the amount of corticosterone in their blood plasma was determined by ELISA (ELISA kit, Abcam), and an open field test was also performed, and the weight of the adrenal glands and thymus was measured. All parameters were compared with the control group (n=10), p<0.05 was considered statistically significant. To assess the stress state in rat pups, blood was taken from the tail vein and the amount of corticosterone was determined. Blood was taken at breakfast and examined in the laboratory. The Open Field Test (OFT) condition was observed, in which stressed rat pups showed the following: an attempt to escape from the center of the cage, reduced movement, and sitting in the corners of the cage.The tissues were fixed in 10% neutral formalin solution for 24 hours, placed in paraffin blocks, and 5 μm thick sections were prepared using a microkeratome. Sections were stained with hematoxylin-eosin (to assess the general histological structure, tissue and nuclear-cytoplasmic ratio) and Masson trichrome (to determine the density of connective tissue and collagen fibers). In order to confirm the correctness of the stress model, the amount of corticosterone in the blood of maternal and infant rats in the control and experimental groups was studied. In maternal rats in the control group, the amount of corticosterone in the initial days of the experiment was 122 ng/ml, on days 14-16 after pregnancy it was 227 ng/ml, and on the last days of pregnancy (days 19-21) it was 546 ng/ml. In the mother rats of the experimental group, this indicator was equal to the higher indicators compared to the control group in the first days of the experiment, it was 160 ng/ml in the first days of the experiment, 454 ng/ml in the 14-16th day of pregnancy and 1165 ng/ml in the last days of pregnancy (Figure 1). 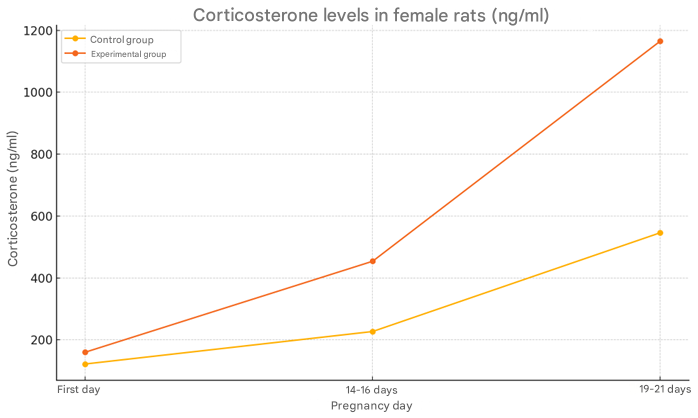 | Figure 1. Changes in the amount of corticosterone in the blood of mother rats in control and experimental groups |
The level of corticosterone in baby rats in the control group changed as follows: 13 ng/ml at 3 days, 18 ng/ml at 7 days, 39 ng/ml at 14 days, 65 ng/ml at 21 days, and 85 ng/ml at 28 days. In the rats of the experimental group, the level of corticosterone changed as follows: at 3 days, it was 25 ng/ml, at 7 days it was 34 ng/ml, at 14 days it was 68 ng/ml, at 21 days it was 100 ng/ml, and at 28 days it was 145 ng/ml (Figure 2).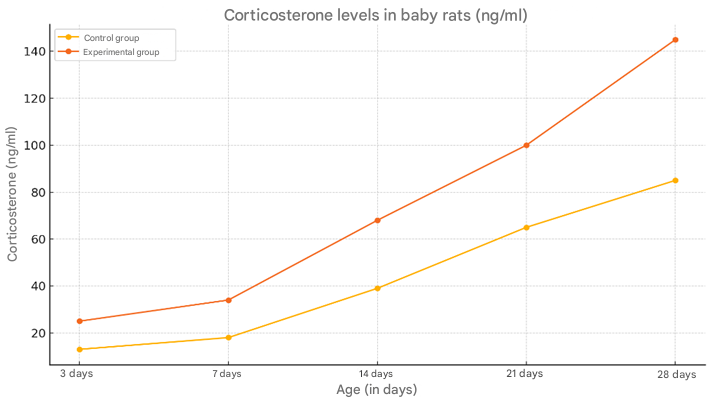 | Figure 2. Changes in the amount of corticosterone in the blood of young rats in the control and experimental groups |
It was observed that the body weight of offspring born to mothers in a state of stress was smaller than that of the control group. By 30 days, it was found that the body weight of young rats in the experimental group was also lower than that of the control group. The body weight of young rats in the control group was 6.1±0.26 grams on average, while in the experimental group, the body weight was 18% less and averaged 5.03±0.23 grams. In the control group, the body weight of 7-day-old rat pups increased by 66.6% compared to the body weight of newborns and averaged 18.5±0.32 grams, while in the experimental group, a 20.5% growth lag was observed compared to the control group and the body weight averaged 14.75±0.37 grams (Figure 3).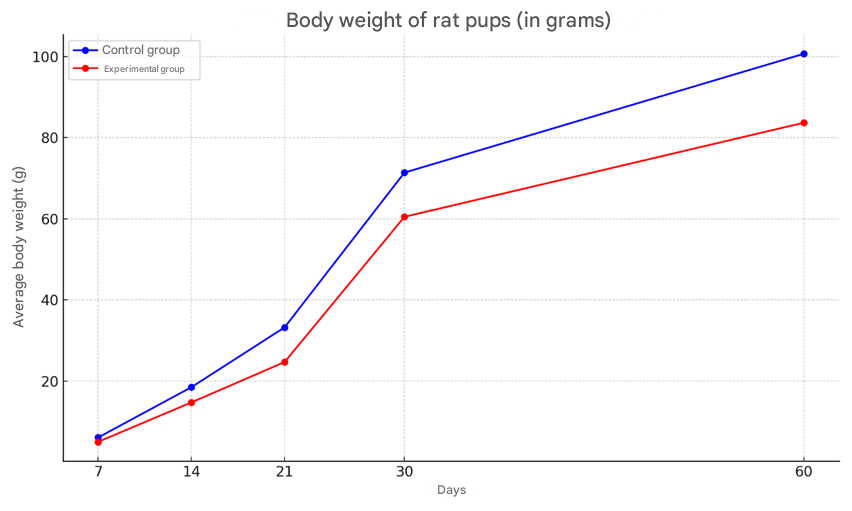 | Figure 3. Body weight changes of rat pups in the control and experimental groups |
The body weight of 14-day-old rat pups born to control rats averaged 33.25 ± 0.44 grams, with a growth rate of 44.1%. The body weight of the offspring in the experimental group during this period was found to lag behind in growth by the most significant (by 25.6%) and averaged 24.7 ± 0.23 grams. In the control group, the body weight of 21-day-old rat pups was observed to increase significantly by 31.4% compared to the body weight of the offspring in the previous group and averaged 48.5 ± 0.88 grams, while in the experimental group, a 17.5% growth lag was observed compared to the control group and averaged 40 ± 1.38 grams.The average body weight of 30-day-old rat pups born to control rats was 71.4±0.97 grams, with a growth rate of 32%. The body weight of the offspring in the experimental group during this period was 60.5±0.63 grams, which was 15.2% less than that in the control group. By the 60-day period, the body weight of the rats in the control group had increased by 29% compared to the previous period, with a body weight of 100.7±2.4 grams. The body weight of the rats in the experimental group was 83.7±1.0 grams, which was 16% less than that in the control group. When the histological structure of the brains of the offspring born to mothers in the experimental and control groups was compared, it was found that by 14 days the cortex was fully formed. In the experimental group of rats, as in the control group, 2-4 nuclear bipolar cells are detected in the cerebral cortex. The 2nd and 3rd layers of the cerebral cortex are densely located. At this stage, the difference in the cerebral cortex of the control and experimental groups is not noticeable. The cortical neurocytes in the experimental group are relatively round in shape and smaller in size than in the control group. These neurocytes have a small rim in their cytoplasm, and the nucleus is brightly colored and relatively larger in size. In control animals, the basophilic substance is formed in the form of flakes in the apical and basal zones of the cytoplasm in large cells, while in experimental animals the cytoplasm of such cells is diffusely stained with thionin, and Nissl's substance is located in the perinuclear area of the cytoplasm.The brains of the offspring born from the control group of rats were fully formed by day 21, the morphological structure of neurocytes was fully formed. In most of the large and small cells of various sizes in different layers of the cortex, basal growths were clearly visible, and the nuclei were mainly round in shape.In the brains of the offspring of the experimental group of 21 days, neurocytes were pale in color, the cytoplasm was not well developed, round in shape, neurocytes were located perinuclearly in most cases, and in some neurocytes they were located in the basal part. Neuroocytes had a rough structure, and the boundaries of the nuclei were clear. By this period of the experiment, a decrease in neurocytes was observed. It was found that the sizes of neurocytes were smaller than those in the control group on this day of the experiment. This was especially clearly reflected in the cytoplasm. The boundaries of the cytoplasm and nucleus were not clear when compared with the control group. The location of the basophilic substance in the cytoplasm of neurocytes located in the hippocampus differed from that in the control group. It was found that the basophilic substance is located in the center of the cytoplasm, and the periphery is more fluid. By the 21st day of the experiment, pathologically changed cells began to be detected in the hippocampus. Here, one can find single cells with a whitish cytoplasm and swollen nuclei.By the first days of postnatal ontogenesis, it was found that most of the large and small neurocytes in the experimental offspring were clearly vacuolated. In this case, perinuclear swelling in the cytoplasm, large and small vacuoles located on the periphery, or neurocytes filling the entire cytoplasm, can be found. The neurocytes in the experimental group have a round shape, and their cytoplasm resembles a mesh. The nuclei of these neurocytes are surrounded by a mesh-like substance like a rim. It was found that the nuclei of these cells are often swollen. The nuclei are fluid, in some cases basophilic. During this period of the experiment, the number of gliocytes was found to be 22% lower than in the control group. The number of perineuronal satellites was 30% lower than in the control group.By the 30th day of the experiment, it was found that large and medium-sized neurocytes in the hippocampus were vacuolated, and perinuclear swelling was present in their cytoplasm. The boundaries of the nuclei of these cells were clearly defined, and the nucleus was visible as a small black dot. During this period, shadow cells of neurocytes were detected. During this period, focal lysis of neurocytes was detected (Figure 4).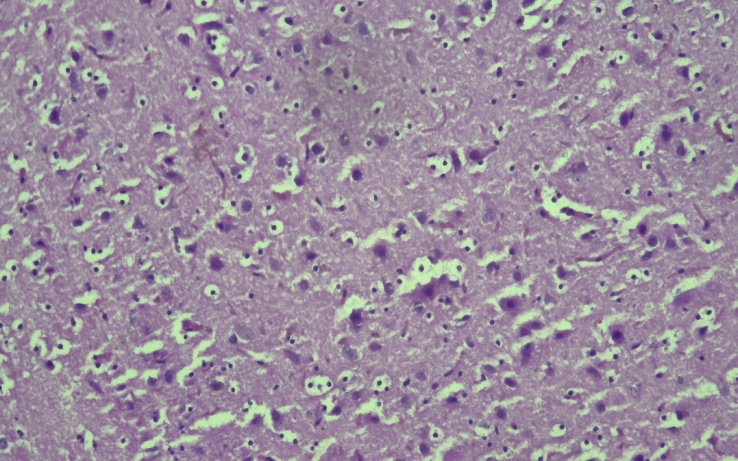 | Figure 4. Appearance of the hippocampus of the experimental group of 30-day-old rats. It was found that the large and medium-sized neurocytes of the hippocampus were vacuolated, and perinuclear edema was present in their cytoplasm. Stain G.E. Dimensions 40x10 |
By the 60th day of the experiment, the structure of the neurocytes in the experimental group was different from that in the control group. By this period, cells with vacuolated cytoplasm were found singly, and the cytoplasm of the neurocytes mainly had a reticular structure. Basophilic substance was located around the nucleus. The nuclei of these cells were found to be of a light color.Their size was increased compared to that in the control group. Chromatin balls were found to be located in most cases on the periphery of the nucleus.According to the results of the experiment, it was found that in children born to rats exposed to prenatal stress, the number of GFAP-positive astroglial cells in the CA1 area of the hippocampus was significantly higher from the 7th day of life compared to the control group. In particular, in the stress group, this indicator averaged 300–320 cells/mm², while in the control group it was around 220–230 cells/mm². These differences were also preserved on days 14, 21 and 28. The largest difference was noted on day 14: 420–450 cells/mm² in the stress group, and 300 cells/mm² in the control group (an increase of more than 40% in the experimental group). These changes are mainly explained by the activation of the hypothalamic-pituitary-adrenal system in the fetus and an increase in the level of glucocorticoids (especially corticosterone). Changes in the proliferation and differentiation of glial cells occur under the influence of long-term stress. The nucleus-cytoplasmic ratio in hippocampal neurons was maintained in normal anatomical proportions in the control infant rats at 3, 7, 14, 21, and 28 days of age. In the early postnatal period (3–7 days), the cytoplasm of neurons is relatively underdeveloped, and the size of the nucleus dominates. But on days 14–28, due to the expansion of the cytoplasm, the ratio decreases slightly, and the nucleus/cytoplasm ratio is around 0.75–0.85, which corresponds to the physiological development process.Significant changes in the nucleus-cytoplasmic ratio were observed in the rat children in the experimental group. In the experimental group, in the hippocampus of 3-day-old rat children, cytoplasm was significantly less than the size of the nucleus, and this ratio was found to be 0.92–0.95. This showed that the development of the cytoplasm was slowed down. The nucleus/cytoplasm ratio was kept around 0.88–0.90 because the growth of cytoplasm in the hippocampal cells of 7–14-day-old rats in the experimental group was slower than that of the control group. At 21–28 days, due to the delay in the morphological maturation of hippocampal neurons, the cytoplasm relative to the size of the nucleus did not increase sufficiently, the ratio remained around 0.85 (in the control group, it decreased to 0.75 at this time).In offspring born to mothers in a state of stress, the cytoplasmic development of hippocampal neurons is impaired, which is expressed in a high nuclear-cytoplasmic ratio. This indicates a decrease in neuronal metabolic activity and the presence of functional defects.Thus, the results of this study showed that offspring born to mothers in a state of chronic stress undergo morphological changes in the hippocampus. When comparing the body weight of rat pups in the experimental and control groups, neurocyte structure and pathomorphological changes in the hippocampus, negative effects were observed in the offspring exposed to stress. The body weight of rat pups in the experimental group was lower than that of the control group, and their growth rate was found to be lower. Also, a decrease in neurocytes, vacuolization in the cytoplasm, perinuclear edema and nuclear changes were noted in their hippocampal tissue. The lysis of neurocytes and the appearance of pathologically altered cells indicate that neurons in the hippocampus can be damaged as a result of stress. All this confirms that stress has a serious impact not only on mental health, but also on brain structure and cognitive functions. Therefore, it is important to develop specific measures to prevent and reduce the effects of chronic stress, to continue research on protecting the hippocampus and supporting cognitive functions.
References
| [1] | Hamilton, G.F., Boschen, K.E., Goodlett, C.R., Greenough, W.T. and Klintsova, A.Y., 2016. Behavioral deficits induced by third-trimester equivalent alcohol exposure in male C57BL/6J mice are not associated with reduced adult hippocampal neurogenesis but are still rescued with voluntary exercise. Behavioural Brain Research, 314, pp. 96–105. https://doi.org/10.1016/j.bbr.2016.07.052. |
| [2] | Dubek, J., Lebel, C., Righini, A., Frayne, R., Beaulieu, C. and Reynolds, J.N., 2014. Hippocampal abnormalities in youth with alcohol-related neurodevelopmental disorder. Journal of the International Neuropsychological Society, 20(2), pp. 181–191. |
| [3] | Alizadeh, A.A., Eisen, M.B., Davis, R.E. et al., 2000. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature, 403(6769), pp. 503–511. https://doi.org/10.1038/35000501. |
| [4] | Perou, C.M., Sorlie, T., Eisen, M.B. et al., 2000. Molecular portraits of human breast tumours. Nature, 406(6797), pp. 747–752. https://doi.org/10.1038/35021093. |
| [5] | Haibe-Kains, B., Culhane, A.C., Desmedt, C., Bontempi, G., Quackenbush, J. and Sotiriou, C., 2010. Robustness of breast cancer molecular subtypes identification. Annals of Oncology, 21(Suppl 4), pp. iv49–iv59. |
| [6] | Liedtke, C., Hatzis, C., Symmans, W.F. et al., 2009. Genomic grade index is associated with response to chemotherapy in patients with breast cancer. Journal of Clinical Oncology, 27(19), pp. 3185–3191. https://doi.org/10.1200/JCO.2008.18.5934. |
| [7] | Teschendorff, A.E. and Caldas, C., 2009. The breast cancer somatic ‘muta-ome’: tackling the complexity. Breast Cancer Research, 11(2), p.301. https://doi.org/10.1186/bcr2236. |
| [8] | World Health Organization (WHO), 2021. Stress at the workplace. [online] Available at: https://www.who.int/news-room/questions-and-answers/item/stress [Accessed 23 Jul. 2025]. |
| [9] | McEwen, B.S., 2007. Physiology and neurobiology of stress and adaptation: central role of the brain. Physiological Reviews, 87(3), pp. 873–904. https://doi.org/10.1152/physrev.00041.2006. |
| [10] | Squire, L.R., 2009. Memory and brain systems: 1969–2009. Journal of Neuroscience, 29(41), pp. 12711–12716. https://doi.org/10.1523/JNEUROSCI.4078-09.2009. |

 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML