-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
American Journal of Biochemistry
p-ISSN: 2163-3010 e-ISSN: 2163-3029
2017; 7(2): 27-36
doi:10.5923/j.ajb.20170702.03

Comparative Study on Fungal Pretreatment and Hydrolysis of Cassava Peelings and Rice Husks for Second-Generation Bioethanol Production
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTMLMichael N. Worfa1, Samson P. Salifu1, Benjamin Afotey2, Moses Mensah2
1Department of Biochemistry and Biotechnology, College of Science, Kwame Nkrumah University of Science and Technology, Ghana
2Department of Chemical Engineering, College of Engineering, Kwame Nkrumah University of Science and Technology, Ghana
Correspondence to: Benjamin Afotey, Department of Chemical Engineering, College of Engineering, Kwame Nkrumah University of Science and Technology, Ghana.
| Email: |  |
Copyright © 2017 Scientific & Academic Publishing. All Rights Reserved.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/

Fluctuating oil prices and its increasing environmental concerns have revived widespread interest in production of biofuel from renewable (lignocellulose) materials. Rice husk and cassava peelings (agro-wastes with little or negligible values to industries in Ghana) were evaluated as a substitute cost effective feed stock for bioethanol production. This project investigated second-generation bioethanol production by pretreating and hydrolysing agro-waste using Pleurotus ostreatus, Aspergillus niger and a combination of the two fungi. The various hydrolysates obtained were subsequently fermented to ethanol using Saccharomyces cerevisiae. The analysis of lignocellulose fractions was conducted using van Soest refractometer whilst fermentable sugars and bioethanol produced were analysed using gravimetric method. The combination of the fungi gave a better yield of fermentable sugars compared to the yield obtained from hydrolysis by either P. ostreatus or A. niger. Of the two fungi, P. ostreatus hydrolysis of rice husk and cassava peelings gave optimum fermentable sugar concentrations of 2.0 g/L and 34.11 g/L respectively, which were higher than 1.33 g/L and 28.64 g/L obtained from A. niger hydrolysis of rice husk and cassava peelings, respectively. The combination of the two fungi for hydrolysis gave the best results for fermentable sugar of 3.0 g/L and 36.51 g/L for rice husk and cassava peelings respectively, for equal weights of the two substrates. The fermentations results revealed that the maximum ethanol yields for cassava peelings and rice husk were 19.36% and 1.53% (w/w dry biomass), respectively. Hence, it can be concluded that cassava peelings can serve as a better feedstock for production of second-generation bioethanol.
Keywords: Second-generation bioethanol, Lignocellulose, Fermentation, Pleurotus ostreatus, Aspergillus niger and Saccharomyces cerevisiae
Cite this paper: Michael N. Worfa, Samson P. Salifu, Benjamin Afotey, Moses Mensah, Comparative Study on Fungal Pretreatment and Hydrolysis of Cassava Peelings and Rice Husks for Second-Generation Bioethanol Production, American Journal of Biochemistry, Vol. 7 No. 2, 2017, pp. 27-36. doi: 10.5923/j.ajb.20170702.03.
Article Outline
1. Introduction
- An alcohol made by fermentation, mostly from carbohydrates produced in sugar or starch crops is termed as bioethanol. Bioethanol is a product of microbial fermentation as opposed to synthetic ethanol produced from petrochemical sources. The fermentation of glucose or other sugars in biomass produces ethanol and carbon dioxide expressed as:
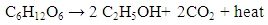 | (1) |
2. Materials and Method
2.1. Sources of Experimental Materials
- The cassava peelings were obtained from cassava dumpsite in Kumasi, Ghana. The rice husks were obtained from a rice farm at Akuse in the Greater Accra Region of Ghana. Spawn of P. ostreatus k910 used for biological pretreatment/enzymatic hydrolysis was obtained from Robart Farms, Kumasi. A. niger Mena1E was also used for biological pretreatment and enzymatic hydrolysis and was obtained from the Pathology Laboratory of the Faculty of Agriculture, Kwame Nkrumah University of Science and Technology, Kumasi. Baker’s yeast used for ethanol fermentation was acquired from Saf-instant, Wisconsin, USA.
2.2. Preparation of A. niger Spore Suspension for Hydrolysis
- Under aseptic conditions, 20 mL of the sterile potato dextrose agar (PDA) acquired from Oxoid (CM1055) Fischer Scientific, Waltham MA, U.S.A was poured into sterilized petri dishes and inoculated with pure cultures of A. niger. The A. niger was subjected to lactophenol cotton blue staining, which confirmed the morphology and spore colouration characteristic of the species. A sterile inoculation pin was used to pick the spores from the pure culture unto the PDA in eight replicates. The inoculated Petri plates were incubated at room temperature for four days after which spores of the mold were collected by flooding the cultures with 20 mL sterile distilled water to dislodge the pores. The spore suspensions were then transferred into a sterile 250 mL Erlenmeyer flask and kept for further use. Serial dilutions of 1/10,000 mL were prepared. The diluted suspension was mixed thoroughly, a drop was sampled with a pipette, and the cell population was determined using a haemocytometer and light microscope.
2.3. Preparation of Stock Culture of S. cerevisiae for Ethanol Fermentation
- Under aseptic condition, 2 g of dry baker’s yeast was transferred into 10 mL of sterile nutrient broth in a McCartney bottle. The bottle was shaken rigorously for even distribution of the cells and incubated for 48 h for activation. Aseptically, an aliquot (0.1 mL) of the cell suspension was then used to inoculate yeast extract agar and the plates incubated at 30°C for 48 h. Pure colonies were subcultured on yeast extract agar and the pure culture was maintained on agar slants at 4°C [8]. Pure culture of a single colony of the yeast growing on the yeast extract agar plate was subsequently inoculated into 50 mL of sterile nutrient broth in a 100 mL Erlenmeyer flask and incubated at 30°C for 48 h in a shaking incubator at 100 rpm. This was used for the ethanol fermentation. The cell population was counted with the aid of a haemocytometer and light microscope. In addition, an aliquot of 1 mL of the stock culture was pipetted into 9 mL of the sterile peptone water in a McCartney tube. Serial dilutions up to fivefold were prepared and finally, 100 μL of each serial dilution was plated on malt extract agar and incubated at 30°C for 24 h. The viable cells were counted and colony-forming units determined.
2.4. Substrate Preparation
- Dried cassava peelings and rice husks, which served as substrates for the hydrolysis, were ground using a blender and sieved with 2 mm sized mesh to obtain a uniform particle size. Quantities of 50 g of the various substrates were weighed into 250 mL conical flasks containing 50 mL of distilled water. With the aid of a spatula the contents were thoroughly mixed. This was done to obtain the optimum moisture content for the inoculum finally resulting in a weight of 100g. The flasks were then corked with absorbent cotton wool and covered with aluminium foil. These were done in triplicate for eight weeks. The conical flasks with their contents were then autoclaved at 121°C at 15 psi for 15 min.
2.4.1. Preliminary Study
- In order to determine the sequence of hydrolysing the substrates with fungi in the double hydrolysis stage, a preliminary study was conducted. With the aid of a blender, 50 g of cassava peelings were milled into 2 mm particle size and mixed uniformly with 50 mL of distilled water. This was then transferred into a conical flask after the pH was determined and the flask corked and autoclaved. After cooling, the substrate in each conical flask was inoculated with either P. ostreatus spawn, or A. niger spore suspension independently in triplicate and incubated for a one-week period at room temperature. Growths were recorded in both fungi-inoculated substrates after the one-week incubation period. The P. ostreatus and the A. niger separately hydrolysed cassava peelings were autoclaved. The autoclaved P. ostreatus hydrolysed cassava peelings were then inoculated with the A. niger spore suspension. The autoclaved A. niger hydrolysed cassava peelings were also inoculated with P. ostreatus spawn and incubated at room temperature for another period of seven days. This was done in triplicate. The concentrations of sugar produced by the various hydrolysed substrates with the fungi were not determined at this stage. At this stage, the main purpose was to determine the sequence of inoculating the inoculum.
2.5. Determination of Moisture Content of Unhydrolysed Samples
- Quantities of 2 g of the untreated but autoclaved samples were transferred into a cleaned crucible (weight known) in triplicate. The individual crucibles with their contents were put in an oven at a temperature of 110°C for 24 h. At the end of the 24 h, the crucibles were taken from the oven, allowed to cool in a desiccator at room temperature and reweighed. Moisture content was calculated by difference and expressed as a percentage of the initial weight of the samples [9].
2.6. Biological Pretreatment and Enzymatic Hydrolysis of the Substrates with P. ostreatus
- After autoclaving, the substrates were allowed to cool at room temperature. Under aseptic conditions, the bottle containing the spawns of P. ostreatus was shaken to loosen the grains. Ten grams of the grains was weighed into a sterile container and subsequently evenly spread onto the prepared substrates, thus a ratio of 1:5 in each of the conical flasks and corked with cotton wool [10]. The experiments were done in triplicate for each substrate. In the negative experimental control set up, spawns of P. ostreatus were not placed onto the substrate. The inoculated substrates and the control were incubated at 28°C for a period of eight weeks with weekly interval terminations of cultures for each substrate. The termination of the culture was to stop the organism from further hydrolysing the substrate. This was done by flooding it with sterile distilled water to the extract released sugars. Aliquots of the hydrolysates of each substrate were then taken for further analysis.
2.7. Biological Pretreatment and Enzymatic Hydrolysis of the Substrates with A. niger
- After autoclaving, the substrates were allowed to cool to room temperature. Under aseptic conditions, 10 mL of the spore suspension (with a concentration of 2.2 x 106 cells / mL) was poured unto the substrates in each flask and corked again. The above spore concentration was used because a minimum level of inoculum was required for effective colonisation and subsequent delignification. A further increase in the inoculum level had insignificant effect on fungal colonisation rate and subsequent growth [11]. The experiments were done in triplicate for each substrate. A negative control experiment was set up in which the substrates were not inoculated with the A. niger suspensions. The inoculated substrates and the control were incubated at 28°C for a period of eight weeks with weekly interval terminations of cultures. The termination of the culture was to stop the organism from further hydrolysing the substrate. This was done by flooding it with sterile distilled water to the extract released sugars. Aliquots of the hydrolysates of each substrate were then taken for further analysis.
2.8. Biological Pretreatment and Enzymatic Hydrolysis of P. ostreatus Hydrolysed Substrates with A. niger
- The substrates were first hydrolysed with P. ostreatus and the hydrolysis process terminated weekly (for the eight weeks’ period) by autoclaving. These served as substrates for the second stage hydrolysis using the A. niger. Under aseptic conditions, 10 mL of the A. niger spore suspension was poured onto the substrates in each flask and corked. The experiments were done in triplicates for each substrate. In the negative experimental control set up, spawns of P. ostreatus and suspensions of A. niger were not added on the substrate. These were subsequently incubated at 28°C for five days. The incubation period was kept as a constant since it would be used for comparative analysis.
2.9. Extraction of Fermentable Sugars
- At the end of each week, the hydrolysis process was terminated by flooding the cultures with sterile distilled water to immediately extract the sugars. The fermentable sugars from the hydrolysed substrates were extracted by pouring 300 ml of distilled sterile water at 28°C onto the content of the flasks. Afterwards, the flasks were fixed on a rotatory shaker at 200 rpm for 30 min. The mixtures were subsequently filtered using filter cloth and the filtrate collected into another sterile container. Distilled sterile water of 200 mL at 28°C was used to thoroughly rinse the conical flask, and the rinsed water was also filtered. The filtrate collected was centrifuged at 5,000 rpm for 30 min at 28°C and the supernatant stored in freezer at -20°C until needed for downstream application. The residue was oven-dried at 70°C for 72 h, milled to homogeneity and stored for chemical analysis.
2.9.1. Determination of Fermentable Sugars
- Fermentable sugars were determined using a digital refractometer (HI96801). Distilled water was used to blank the refractometer and then 3 mL of supernatant was drawn and dropped on a groove on the refractometer. The total sugar values were read, converted from Brix to g/L and recorded.
2.10. Fermentation of Hydrolysates to Ethanol
- Ethanol fermentation was conducted using a modified method of Zakpaa et al. [8]. Hydrolysates of 30 mL were dispensed into labelled 100 mL bottle and autoclaved at 121°C for 15 min and allowed to cool. The pH of the hydrolysates was adjusted to 5.5 (using sterile NaOH and HCl) and inoculated with 0.3 mL of the S. cerevisiae suspension (containing 2.9 x 106 cells/mL in sterile nutrient broth). In the control set-up however, the hydrolysates were not inoculated with S. cerevisiae. The bottles were tightly capped with polypropylene caps, sealed with the aid of an adhesive tape and incubated at 30°C at 100 rpm for 5 days in a shaking incubator for fermentation. The concentration of ethanol produced was determined at regular intervals of 12 h for the five days’ fermentation duration using the gravimetric method. Thus, the difference in weight due to carbon dioxide production was determined every 12 h and this corresponds to mass of ethanol produced. The yields of ethanol produced were also determined using a relation; 1. Ethanol Yield (%W/W Sugar) = ((Mass of ethanol produced in reactor (g))/(mass of sugar in hydrolysate (g)) ×0.51)×100, where 0.51 is a constant conversion factor [3]. 2. Ethanol Yield (%W/W Dry Biomass) = ((Mass of ethanol produced in reactor (g))/(mass of dry biomass (g))×0.51)×100, where 0.51 is a constant conversion factor [3].
2.11. Chemical Analysis
- The chemical composition of the raw and the inoculated substrates was determined by proximate analysis using van Soest Method of Analysis [12] for the determination of the fibre components (neutral detergent fibre (NDF), acid detergent fibre (ADF), and acid detergent lignin (ADL)) of the hydrolysed and un-hydrolysed substrates. These were used to estimate the cellulose, hemicellulose and lignin contents. Thus 1. NDF = Cellulose + Hemicellulose + Lignin 2. ADF = Cellulose + Lignin 3. Hemicellulose = NDF – ADF 4. Cellulose = NDF– Hemicellulose –Lignin5. Lignin = ADF – CelluloseThe means ± SD for each of the three replicates in every experiment were determined. Using a significance level of p < 0.05, an analysis of variance (ANOVA) was carried out and graphs (with standard deviation errors bars) plotted using Graph Pad Prism software (Version 6.0).
3. Results
3.1. Preliminary Study Results
- It was observed that P. ostreatus was unable to colonise and hydrolyse both the rice husk and cassava peelings substrates that were previously hydrolysed with the A. niger. However, A. niger was able to colonise and hydrolyse both substrates that were previously hydrolysed by P. ostreatus under the same conditions as the latter. This was used to determine the sequence of hydrolysis in the two-stage hydrolysis process.
3.2. Moisture Content of the Unhydrolysed Samples
- The determination of the moisture content of unhydrolysed but autoclaved rice husks and cassava peelings samples gave moisture content results of 70.82% and 70.26%, respectively. These served as the optimum moisture content for the growth of the microorganism being used for the initial hydrolysis. During the moisture content determination, a desiccator was used to ensure no moisture was absorbed.
4. Discussion
- Preliminary studies were carried out to determine the sequence of hydrolysis in the two-stage fungal hydrolysis since growth of fungi could be inhibited by another fungus. The preliminary studies revealed that P. ostreatus was unable to colonise and hydrolyse both the rice husks and cassava peelings substrates that were previously hydrolysed with the A. niger. This could be a result of secondary metabolites such as aflatoxins, patulins and satratoxins produced by A. niger [13] which could be inhibitory to the colonisation of P. ostreatus. However, A. niger was able to colonise and hydrolyse both substrates that were previously hydrolysed by P. ostreatus under the same conditions as the latter, suggesting no inhibitory metabolites were produced by P. ostreatus. Bentil et al. [10], reported similar observations in their work. The preliminary studies results informed the decision to either hydrolyse the pretreated cassava peelings and the rice husks with A. niger alone or to first hydrolyse the pretreated cassava peelings and the rice husks with P. ostreatus and then followed by A. niger.There was no fermentable sugar released immediately after autoclaving the rice husks. This could be due to the fact that rice husk is low in starch content, but has very high recalcitrant lignin, which strongly shields the cellulose and hemicellulose from degradation to monomer sugars units by autoclaving [14]. In contrast, although the initial fermentable sugar concentrations of the raw substrates were not determined before autoclaving, the sugar concentrations of the cassava peelings immediately after autoclaving were very high. This could be due to the fact that since cassava peelings have high starch content, subjecting it to autoclaving caused the release of monomer sugar units [15] which reflected in the graphs for the controls in Figures 4, 5 and 6. Thus autoclaving for sterilization of cassava peelings substrates before fungal hydrolysis had an impact on the substrates, resulting in the release of fermentable sugars. It is also possible that some amount of the cassava tuber content, which is rich in monosaccharaides was left attached to the cassava peels surface. This also, could have contributed to the high amount of fermentable sugars present in the cassava peelings hydrolysates after autoclaving. The hydrolyses of cassava peelings and the rice husks with either A. niger or P. ostreatus alone, and the combination of first, with P. ostreatus and then with A. niger resulted in significant increase in fermentable sugars. The optimum fermentable sugars obtained from cassava peelings hydrolysates were significantly higher (p < 0.05) than those of rice husks hydrolysates. This may be due to the low lignin content but high starch content of cassava peelings as compared to rice husk (Figure 1 to 6). The celluloses (and starch) of cassava peelings are readily available and easily hydrolysed into sugar units by the extracellular enzymes produced by fungi as compared to those of rice husks. Cassava peelings treated with P. ostreatus produced highest concentration of 34.11 g/L of sugar as compared to 28.64 g/L obtained with the use of A. niger within one week of hydrolysis (Figure 4 and 15). This may be due to the synergistic nature of cellulases produced by the microorganisms and an indication that the enzymes from P. ostreatus may contain more of the components of the cellulase complex than A. niger [10]. P. ostreatus is also one of the fungi found to have an elaborate higher exoglucanase and endoglucanase activities [15].
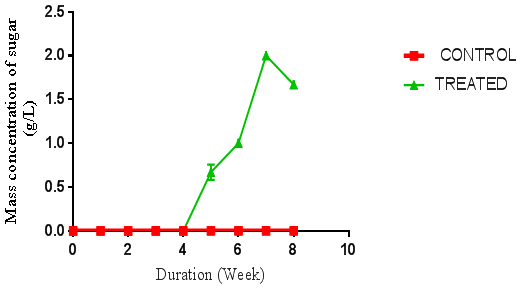 | Figure 1. Concentration of sugar produced from P. ostreatus hydrolysis of rice husk. (Control = unhydrolysed rice husk hydrolysate, Treated = hydrolysed rice husk hydrolysate) using negative control |
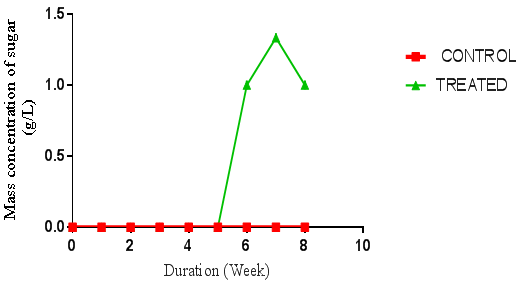 | Figure 2. Concentration of sugar produced from A. niger hydrolysis of rice husk. (Control = unhydrolysed rice husk hydrolysate, Treated = hydrolysed rice husk hydrolysate) using negative control |
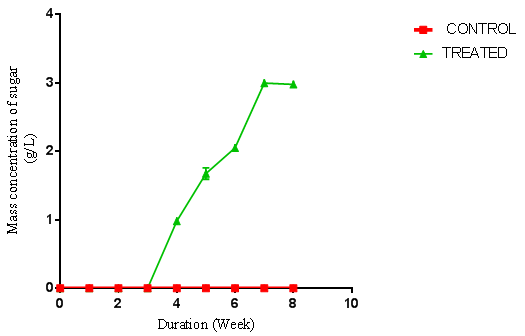 | Figure 3. Concentration of sugar produced from A. niger hydrolysis of rice husk previously hydrolysed with P. ostraetus. (Control = unhydrolysed rice husk hydrolysate, Treated = hydrolysed rice husk hydrolysate) using negative control |
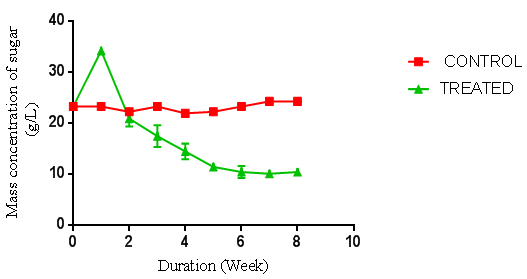 | Figure 4. Concentration of sugar produced from P. ostreatus hydrolysis of cassava peelings. (Control = unhydrolysed cassava peelings hydrolysate, Treated = hydrolysed cassava peelings hydrolysate) using negative control |
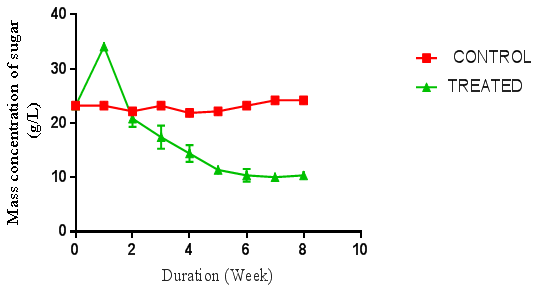 | Figure 5. Concentration of sugar produced from A. niger hydrolysis of cassava peelings. (Control = unhydrolysed cassava peelings hydrolysate, Treated = hydrolysed cassava peelings hydrolysate) using negative control |
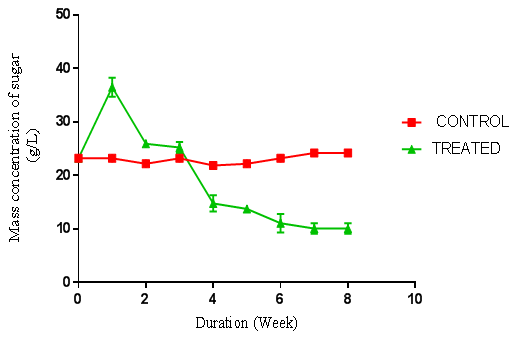 | Figure 6. Concentration of sugar produced from A. niger hydrolysis on cassava peelings previously hydrolysed with P. ostraetus. (Control = unhydrolysed cassava peelings hydrolysate, Treated = hydrolysed cassava peelings hydrolysate) using negative control |
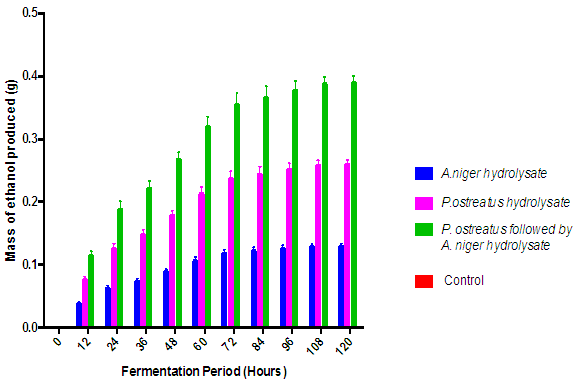 | Figure 7. Ethanol production from the fermentation of the various rice husk hydrolysates with S. cerevisiae (Control = hydrolysate not inoculated with S.cerevisiae) |
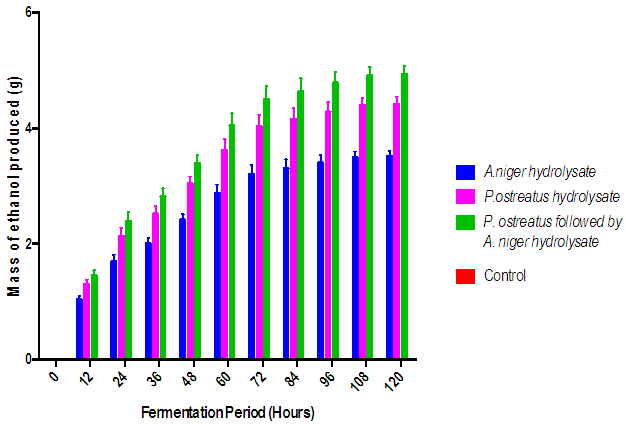 | Figure 8. Effect of fermentation of the various cassava peelings hydrolysate with S. cerevisiae on ethanol production. (Control = hydrolysate not inoculated with S.cerevisiae) |
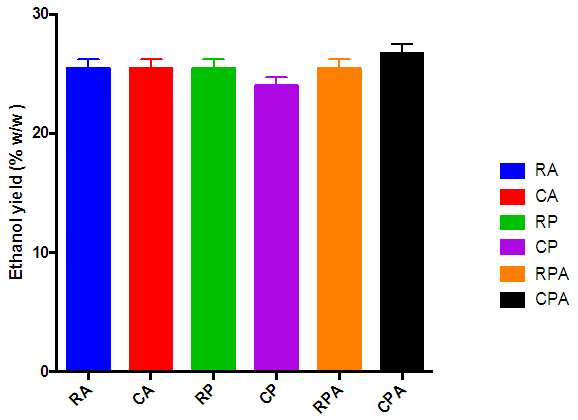 | Figure 9. Percentage Ethanol yield (relative to sugar in the hydrolysate) of the various hydrolysates after 120 h fermentation period. (hydrolysate obtained from hydrolysis of cassava peelings with P. ostreatus followed by A. niger (CPA), hydrolysis of cassava peelings with P. ostreatus(CP), hydrolysis of rice husk with A. niger (RA), hydrolysis of cassava peelings with A. niger (CA), hydrolysis of rice husk with P. ostreatus (RP) and hydrolysis of rice husk with P. ostreatus followed by A. niger (RPA)) |
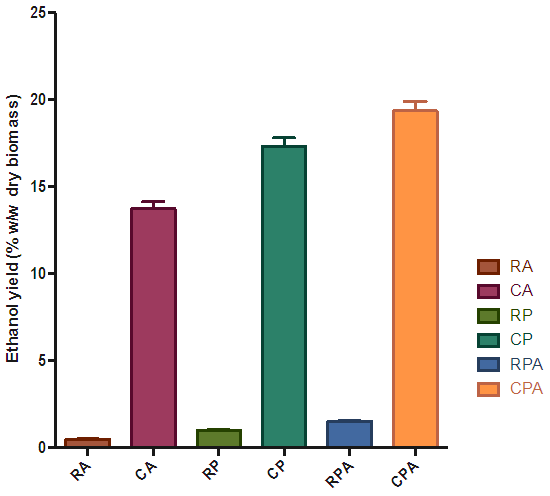 | Figure 10. Effect of type of hydrolysis on percentage ethanol yield of dry biomass |
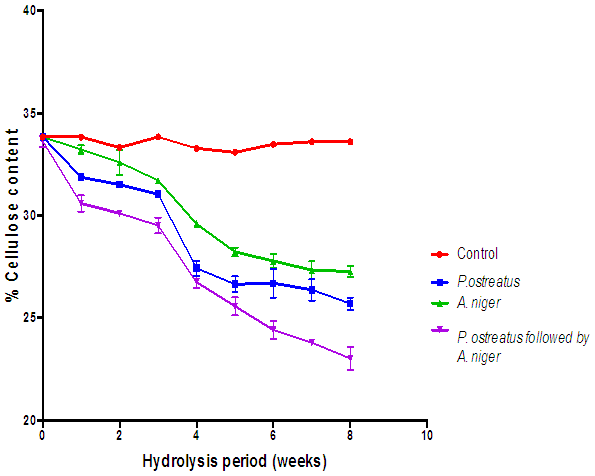 | Figure 11. Comparison of the effect of hydrolysis of rice husk with the various fungi on cellulose content (Control= Rice husks not inoculated with fungi) |
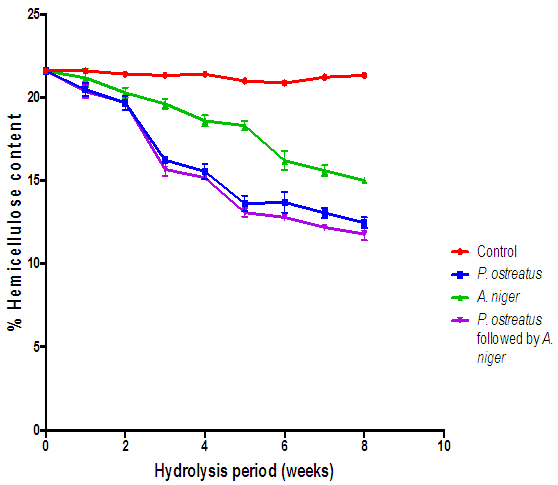 | Figure 12. Compariosn of the effect of hydrolysis of rice husk with the various fungi on hemicellulose content (Control= Rice husks not inoculated with fungi) |
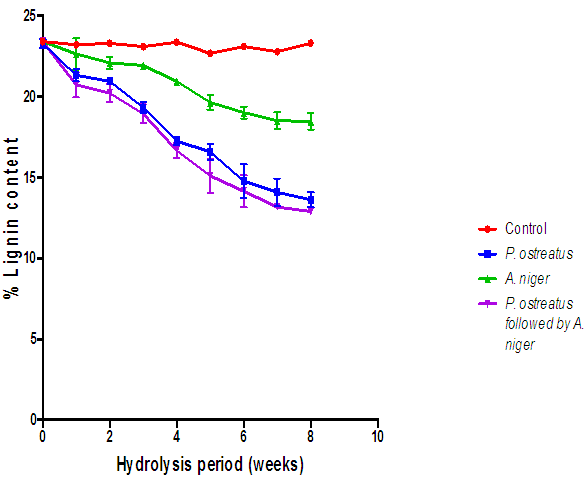 | Figure 13. Comparison of the effect of hydrolysis of rice husk with the various fungi on lignin content (Control=Rice husk not inoculated with fungi) |
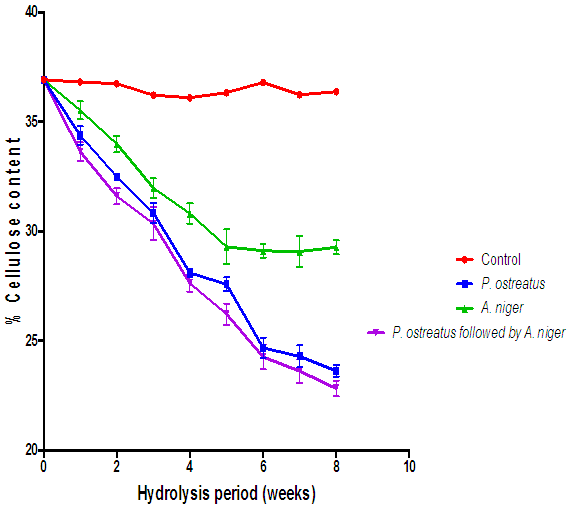 | Figure 14. Comparison of the effect of P. ostreatus and A. niger hydrolysis of cellulose content of cassava peelings (Control=Cassava peelings not inoculated with fungi) |
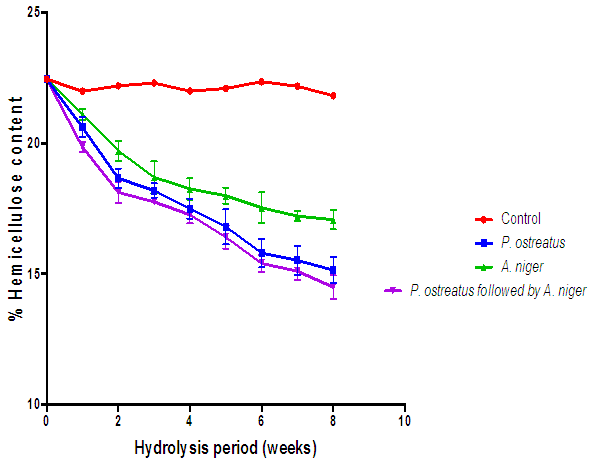 | Figure 15. Comparison of the effect of P. ostreatus and A. niger hydrolysis of hemicellulose content of cassava peelings (Control=Cassava peelings not inoculated with fungi) |
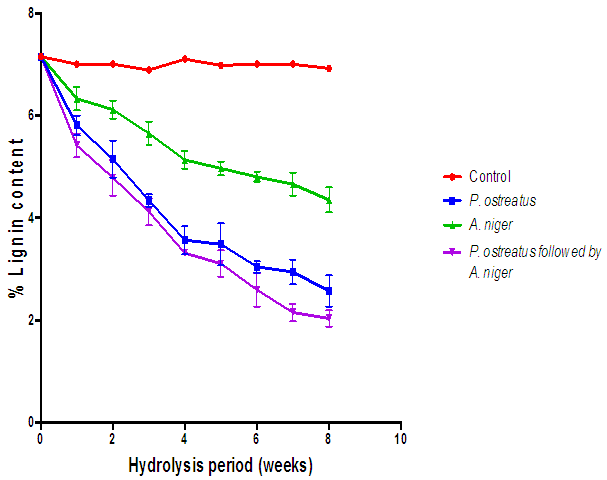 | Figure 16. Comparison of the effect of P. ostreatus and A. niger hydrolysis of lignin content of cassava peelings (Control=Cassava peelings not inoculated with fungi) |
5. Conclusions
- Bioethanol was successfully produced from rice husks and cassava peelings. P. ostreatus, A. niger and the combination of the two fungi were able to hydrolyse the substrates into fermentable sugars. The selected order of the two-stage biological pretreatment/ hydrolysis with P. ostreatus followed by A. niger significantly enhanced the concentration of fermentable sugars produced as compared to the individual fungus. The fermentable sugars produced were converted to bioethanol by S. cerevisiae. Evaluation of the various fermented hydrolysates with S. cerevisiae of the two substrates revealed that cassava peelings produced higher ethanol yield (19.36% w/w dry biomass) than those of rice husks (1.53% w/w dry biomass). These therefore confirmed cassava peelings as a useful and better raw material for second-generation bio-ethanol production.