-
Paper Information
- Paper Submission
-
Journal Information
- About This Journal
- Editorial Board
- Current Issue
- Archive
- Author Guidelines
- Contact Us
American Journal of Biochemistry
p-ISSN: 2163-3010 e-ISSN: 2163-3029
2014; 4(4): 59-67
doi:10.5923/j.ajb.20140404.01
Antidiabetic and Antioxidative Potential of Cystoseira myrica
 Abstract
Abstract Reference
Reference Full-Text PDF
Full-Text PDF Full-text HTML
Full-text HTML1Chemistry department, Faculty of science, Beni-Suef University, Egypt
2Agriculture chemistry department, Faculty of Agriculture, El minia university, Egypt
Correspondence to: Nagy MA, Chemistry department, Faculty of science, Beni-Suef University, Egypt.
| Email: |  |
Copyright © 2014 Scientific & Academic Publishing. All Rights Reserved.
Alloxan administration in male albino rats, induced diabetes by increasing blood glucose concentration and reducing hepatic glycogen content as compared to normal control group. Besides, serum lipid profile parameters such as total-cholesterol, triglyceride and low-density lipoprotein were also elevated, whereas, the level of high-density lipoprotein-cholesterol was reduced significantly (P<0.05) in diabetic rats. Treatment of diabetic animals with hot aquous extract of Cystoseiramyrica for 30 days, significantly lowered blood glucose level, elevated hepatic glycogen content and maintained body weight and lipid-profile parameters towards near normal range. Declined activity of antioxidant enzymes and concentration of non-enzymatic antioxidants were also normalized by drug treatment, thereby reducing the oxidative damage in the tissues of diabetic animals and hence indicating the anti-diabetic and antioxidant efficacy of the extract. Biochemical observations were further substantiated with histological examination of pancreas and liver.
Keywords: Cystoseiramyrica, Diabetesmellitus, Oxidative Stress, Antidiabetic, Antioxidant
Cite this paper: Nagy MA, Ewais MM, Antidiabetic and Antioxidative Potential of Cystoseira myrica, American Journal of Biochemistry, Vol. 4 No. 4, 2014, pp. 59-67. doi: 10.5923/j.ajb.20140404.01.
Article Outline
1. Introduction
- The increasing incidence of diabetes represents an enormous socio-economic burden in the developing countries. The World Heath Organization estimates that over 300 million people worldwide will have Diabetes mellitus (DM) by the year 2025 with alarming proportions from developing countries [1].DM is a chronic disease caused by inherited or acquired deficiency in insulin secretion and by decreased responsiveness of the organs to secrete insulin. There are two main forms of diabetes. Type 1diabetes is due primarily to autoimmune-mediated destruction of pancreatic islet beta-cells, resulting in dramatic insulin deficiency while Type 2 diabetes is characterized by abnormal insulin secretion, associated with varying degrees of insulin resistance [2].Oxidative stress represents a significant mechanism for the destruction of cells that can involve apoptotic cell injury and apoptosis in a variety of cell types that involve endothelial cells (ECs), cardiomyocytes, and smooth muscle cells through multiple cellular pathways. The production of Reactive oxygen species (ROS) can lead to cell injury through a number of processes that involve the peroxidation of cellular membrane lipids [3].Oxidative stress forms the foundation for the induction of multiple cellular pathways that can ultimately lead to both the onset and subsequent complications of DM. Diabetics and experimental animal models exhibit high oxidative stress due to persistent and chronic hyperglycemia, which thereby depletes the activity of antioxidative defense system and thus promotes de novo free radicals generation [4a].The source of oxidative stress is a cascade of ROS leaking from the mitochondria. This process has been associated with the onset of type 1 diabetes (T1DM) via the apoptosis of pancreatic β-cells, and the onset of (T2D) via insulin resistance [5].In recent years due to the adverse effects of synthetic hypoglycemic drugs, interests in alternate therapeutic approach have become very popular Nowadays; herbal drugs are gaining popularity in the treatment of diabetes and its complications due to their efficacy, low incidence of side effects and low cost [6].An increasing number of studies has demonstrated that marine algae is a rich source of compounds which have is a rich source of compounds which have potential therapeutic uses. Brown marine algae to be rich sources of antioxidant compounds with potential free radical scavenging for the prevention and treatment of various diseases caused by oxidative damage. Fucoidans, polysaccharides containing substantial percentages of L-fucose and sulfate ester groups, are constituents of brown algae that have numerous other biological properties such antioxidant, anti-inflammatory immuno-modulatory and apoptosis-inducing activities [7a].Cystoseira myrica (S. G. Gmelin) C. Agardh (Gulf of Suez) is brown marine algae that can be classified as one of the more advanced species (phytochemically) of the Cystoseira genus based on the complexity of the terpenes produced. Hot water extract of C. myrica (CME) is rich in bioactive metabolites derived from algae are as sulfated polysaccharides that have antioxidant activity [8, 9].But till date, It is the first study about the anti-diabetic and antioxidant potential of Cystoseira myrica. In the present study, the effect of 30 day chronic oral treatment with CME with a dose of 400 mg/kg body wt on diabetes and resultant oxidative stress was investigated by evaluating its antihyperglycemic, antihyperlipidemic and antioxidative properties in alloxan induced diabetic rats confirmed by histological examination of pancreas and liver of alloxan induced diabetic rats.
2. Materials and Methods
- Chemicals: Alloxan monohydrate was purchased from Sigma Fine Chemicals. All other chemicals used for this study were of analytical grade and obtained from Stanbio Laboratory USA Kits. Kits for the estimation of total cholesterol, triglyceride and HDL-cholesterol were purchased from Diamond Diagnostic Egypt.
2.1. Preparation of Extract
- Cystoseira myrica was collected from El Zafrana, Gulf of Suez. About 100 g of the fresh alga, corres ponding to 10 – 12 g of dry alga material was homogenized in 500 ml hot double distilled water. The mixture was clarified by filtration using Whatman No.1 filter paper and the light brown extract resulted. The water extract of H. clathratus were sterilized by filtration and autoclaving, respectively.
2.2. Animal Care and Monitoring
- 30 Healthy male albino rats (4-5 months old, weighing 190-210 g) were procured from Faculty of Agricultural, El Minia University, Egypt. They were housed under standard laboratory conditions of light (12:12 h L: D cycle), temperature (23 ± 2°C) and relative humidity (55 ± 5%). The animals were provided standard rat pellet feed and tap water ad libitum. Maintenance and treatment of all the animals was done in accordance with the principles of Institutional Animal Ethics Committee constituted as per the directions of the Committee for the Purpose of Control and Supervision of Experiments on Animals, Egypt.
2.3. Induction of Diabetes and Treatment with Drugs
- For the present study, animals were divided into following 3groups:NC (normal control), DC (diabetic control) and CME (diabetic + Cystoseira myrica extract treated). After fasting for 18 hours, rats of group DC, CME were made diabetic by a single intra-peritoneal injection of alloxan monohydrate, 150 mg/kg body wt, freshly dissolved in normal saline [10]. Subsequent to alloxan administration the rats had free access to food and water and were provided with 50% glucose solution to drink overnight to counter drug induced hypoglycemic shock. One week after alloxan injection, the fasting blood glucose (FBG) concentration was determined by means of one touch ultra glucometer (Johnson & Johnson Company, USA) and compatible blood glucose strips [11]. Rats showing fasting blood level greater than 140 mg/dl were considered diabetic and selected for treatment with hot water extract of Cystoseira myrica (CME) (400 mg/kg body wt.). The CME were administered orally, once in a day for 30 days.
2.4. Biochemical Estimations in Blood and Serum Insulin
- At the end of drug treatment, the animals were fasted over night but allowed free access to water .On the next morning, all groups were bled by vein puncture, fasting blood samples were centrifuged, sera were kept at -80℃ right the time of analysis, Then, rats were sacrificed by decapitation and abdominal incision for separation of pancreas, and liver. After that separated organs was were (g.) kept at -20°C for measurement of oxidative stress profile in homogenate of pancreas and liver. Also, other part of the separated organs was forwarded for histological staining.Fasting blood glucose (FBG) concentration of all the three experimental groups was determined by glucometer during different phases of the experiment by withdrawing blood from the tail vein. Serum insulin was assayed in the Radioactive Isotopes Unit, Central Department of Scinentificial Analysis and Test, National Research Center (Dokki, Giza) by radioimmunoassay kits of DPC (Diagnostic Products Corporation, Los Angeles, USA) [coat-A-count] [12].For estimating serum lipid profile, isolated serum was from overnight fasted rats on day 30th of CME treatment and serum total cholesterol (TC), triglyceride (TG) and HDL-cholesterol were estimated by using diagnostic kits (Erba Mannheim Cholesterol kit, Transasia Bio-Medicals Ltd., Daman) and their results were expressed in mg/dl.
2.5. Biochemical Estimation in Tissue Homogenates
- Pancreas and liver and were removed, freed from adhering tissues and washed with ice-cold normal saline solution (0.9%). Weight of all the organs was taken only after drying the tissue. 1 g tissue was homogenized in 10 ml of 0.2 M tris-HCl with the help of homogenizer. The homogenate was filtered and then centrifuged at 10,000 rpm for 20 minutes at 4°C. The supernatant obtained was used for estimation of superoxide dismutase (SOD) [13], catalase (CAT) [14], glutathione peroxidase (GSH-Px) [15], Reduced glutathione (GSH) [16], and thiobarbituric acid reactive substances (TBARS) [16]. Hepatic G-6-Pase was measured [17] and glycogen phosphorylase was measured [18].
2.6. Histopathological Examination of Tissues
2.6.1. Histopathological Examination of Pancreas
- Some pancreas were cleaned and fixed in 10% neutral buffered formalin solution. After dehydration in graded ethanol solutions and in toluene, they were embedded in paraffin. Sections of 3–5µm thickness were stained with hematoxylin and eosin (H.E.) for histopathological examination.
2.6.2. Histopathological Examination of Liver
- Some liver were cleaned and fixed in 10% neutral buffered formalin solution. After dehydration in graded ethanol solutions and in toluene, they were embedded in paraffin. Sections of 3–5µm thickness were stained with hematoxylin and eosin (H.E.) for histopathological examination. Other sections were stained with (Periodic acid staining) and PAS Stain for histopathological examination of glycogen.
2.7. Statistical Analysis
- Results are expressed as mean ± Standard Error of Mean (SEM). Statistical analysis was performed using one-way Analysis of Variance (ANOVA) followed by Tukey’s post-hoc multiple comparison test using SPSS (version 16.0) and student’s‘t’-test using Sigma Plot (version 8.0). The values of P<0.05 were considered as statistically significant.
3. Results
3.1. Effect on Body Weight
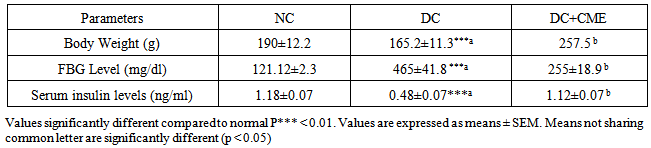
- All animals treated with alloxan in diabetic control group (DC) revealed a significant (P<0.05) loss in body weight, which was persistently observed till the end of the study period (Table 1). Weight reduction induced by diabetes in group (CME) was cured by CME treatment.
|
3.2. Effect on Fasting Blood Glucose Level and Serum Insulin
- Alloxan injection to DC and CME groups resulted in a significant (P<0.05) increase in blood glucose level as compared to normal control group A. Treatment with Cystoseira myrica hot water extract (CME)for 30 days significantly reduced the glucose after 30 days of treatment (Table 1).Alloxan injection to DC and CME groups resulted in a significant (P<0.05) decrease in serum insulin level as compared to normal control group A. Treatment with Cystoseira myrica hot water extract for 30 days significantly enhanced the insulin release after 30 days of treatment (Table 1).
3.3. Effect of CME on Activity of Hepatic Glycogen Phosphorylase and Hepatic Glucose-6- Phosphatase in Alloxan Induced Diabetic Rats
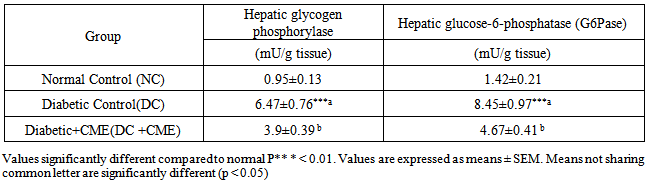
- Activity of hepatic glycogen phosphorylase and hepatic glucose-6- phosphatase increased significantly in diabetic control group (DC) compared to normal control group (NC). However, treatment with Cystoseira myrica hot water extract for 30 days significantly (P<0.05) decreased activity of hepatic glycogen phosphorylase and hepatic glucose-6- phosphatase as compared to DC (Table 2).
3.4. Effect on Serum Lipid Profile
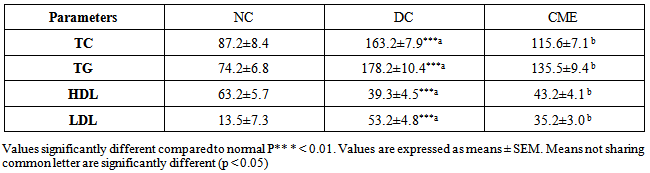
- Compared to normal control group (NC), the level of serum TC, TG, LDL cholesterol was significantly (P<0.05) increased whereas the level of HDL-cholesterol was significantly (P<0.05) reduced by in untreated diabetic rats (DC) (Table 3). After treatment of group CME with Cystoseira myrica hot water extract for 30 days, the level of serum TC, TG, and LDL -cholesterol reduced significantly (P<0.05) whereas, the level of serum HDL-cholesterol was significantly increased.
|
3.5. Antioxidative Status
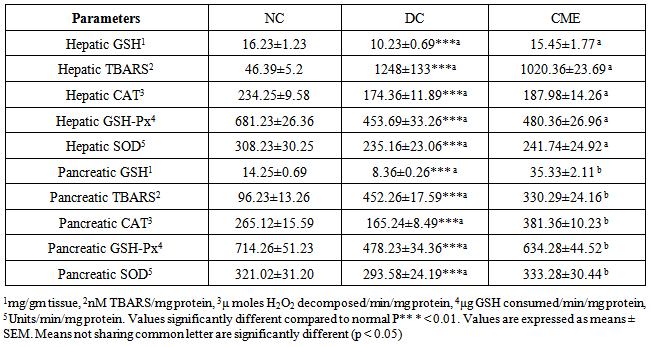
- It was observed that after inducing diabetes, the level of hepatic and pancreatic GSH content decreased significantly in diabetic control group (DC) as compared to normal control group (NC). Treatment of group CME with Cystoseira myrica hot water extract for 30 days significantly (P<0.05) increased the tissue’s GSH level as compared to DC (Table 4).
|
3.6. Effects of CME on Alloxan-Induced Histological Changes in Pancreas
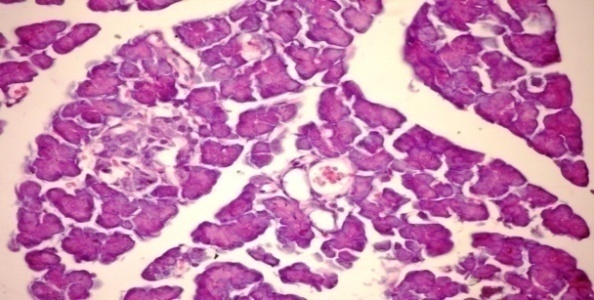
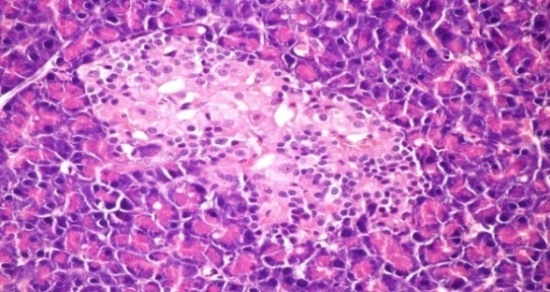
- The results of Cystoseira myrica in histopathologic examination are shown in Fig. 1. As revealed in (Fig. 1b), a clear decrease in the area occupied by the β-cells was observed in the pancreatic sections of alloxan-induced diabetic rats. Treatment with CME showed hypertrophy, hyperplasia and vacuolations of β-cells of islets of langerhans compared to alloxan treated rats (Fig. 1c) revealing the protective effect of CME. Fig (1) Histological changes in pancreases of diabetic rats (H & E):
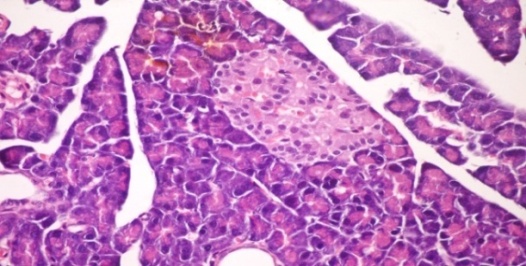 | Figure 1a. Pancreatic tissue of normal male albino rats. The pancreas is subdivided by septa inti pancreatic lobules. The exocrine portion of the pancreas consists of pancreatic acini while endocrine portion consists of islets of Langerhans (H & E x 400) |
 | Figure 1b. Pancreatic tissue of diabetic rats. Normal architecture of the islets is disrupted islets of Langerhans exhibited hydrophobic cells, necrotic cells, vacuolizations and irregular hyperchromic nuclei (H& E x 400) |
 | Figure 1c. Pancreatic tissue of diabetic rats treated with CME (orally 400 mg/kg body weight of rats daily for 30consecutive days) showing hypertrophy, hyperplasia and vacuolations of β- cells of islets of langerhans (H& E x 400) |
3.7. Effects of CME on Alloxan-Induced Histological Changes in liver (H&E Stain)
- The results of Cystoseira myrica in histopathologic examination are shown in Fig. 2. As revealed in (Fig. 2b) kupffer cell activation, dilation and congestion of central vein and hepatic sinusoids were observed in the hepatic sections of alloxan-induced diabetic rats. Treatment with CME showed kupffer cells activation and congestion of central vein compared to alloxan treated rats (Fig. 2c) revealing the hepatotoxic effect of CME.Fig. (2) Histological changes in liver of diabetic rats (H & E):
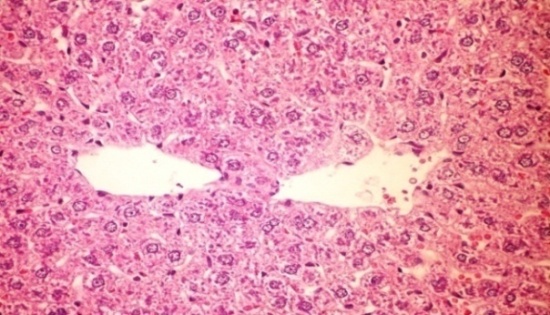 | Figure 2a. Liver of normal control rat showing the normal histological structure of hepatic lobule (H& E x 400) |
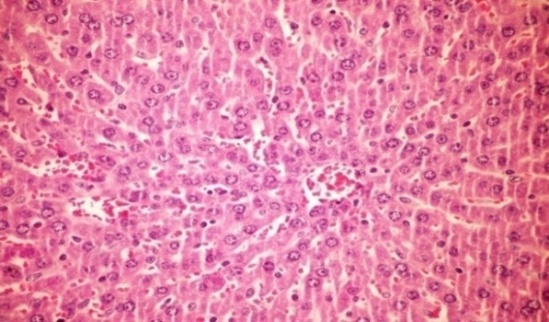 | Figure 2b. Liver of diabetic rat showing kupffer cells activation, dilation and congestion of central vein and hepatic sinusoids (H& E x 400) |
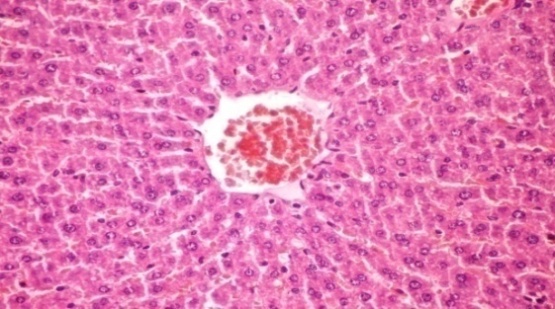 | Figure 2c. Liver section of diabetic rats treated with CME (orally 400 mg/kg body weight of rats daily for 30consecutive days) showing kupffer cells activation and congestion of central vein (H& E x 400) |
3.8. Effects of CME on Alloxan-Induced Histological Changes in Liver (PAS)
- The results of Cystoseira myrica in histopathologic examination are shown in Fig. 3. As revealed in (Fig. 3b), variable sized positive Pas glycogen particles distributed throughout the hepatocytic cytoplasm in the hepatic sections of alloxan-induced diabetic rats. Treatment with CME showed a moderate PAs particles distributed throughout the hepatocytic cytoplasm compared to alloxan treated rats (Fig. 3c).Fig (3) Histological changes in Liver of diabetic rats (PAS):
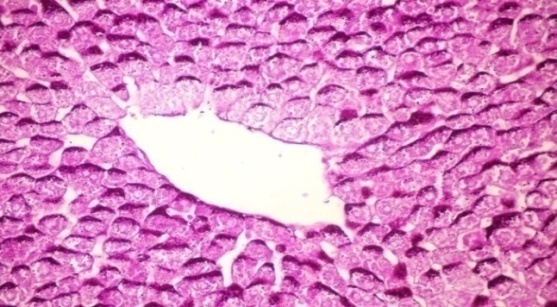 | Figure 3a. Liver of normal control rat showing accumulation of reddish stained glycogen particles at one pole of hepatocytes (PAs stain x 400) |
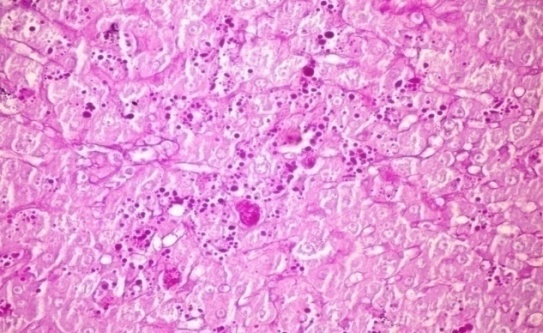 | Figure 3b. Liver of alloxan induced diabetic rat showing variable sized positive Pas glycogen particles distributed throughout the hepatocytic cytoplasm (PAs stain x 400) |
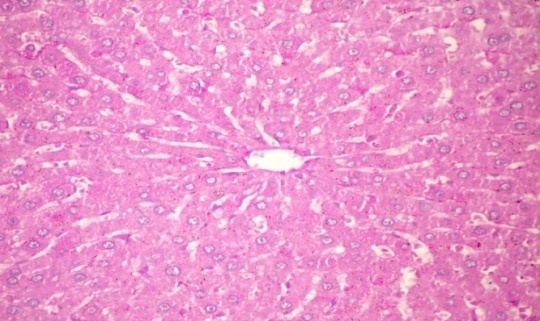 | Figure 3c. Liver of diabetic rats treated with CME (orally 400 mg/kg body weight of rats daily for 30consecutive days) showing no histlogical changes (PAs stain x 400) |
4. Discussions
- Glucose is the key physiological regulator of insulin secretion; indeed, short-term exposure of β-cells to increasing glucose concentrations induces proliferation in a concentration-dependent manner. In addition to its effect on β-cell turnover, hyperglycemia also impairs β-cell secretary function. This glucotoxic effect is evident before apoptosis leads to a significant decrease in β-cell mass [19]. Alloxan –induced diabetic rats exhibited severe glucose intolerance and metabolic stress as well as hyperglycemia due to a progressive oxidative insult interrelated with a decrease in endogenous insulin secretion and release [20]. Treatment with antidiabetic drugs based on their pancreatic antioxidant activity might be a protective strategy for protecting β-cell due to disproportionate generation of free radicals [21, 22].The present investigation indicated that a single dose of alloxan (150 mg/kg) intraperitoneally to adult male albino rats (190-210g) was suitable to induce histological changes of the islets of Langerhans characterized appearance, hypoinsulinemia and hyperglycemic state [23].A gradual loss of β-cells due to apoptosis significantly hinders insulin production and inhibits cell viability. During apoptosis, cells shrink; chromatin condenses; DNA is cleaved into pieces at inter nucleosomal regions. A proactive way to increase β-cell viability is to decrease apoptosis level in order to retain the cell population and increase insulin production [24].Exposure of islets to alloxan shows significantly increased formation of peroxynitrite, NO and ROS with markedly elevated lipid peroxidation and reduced cell viability. Islets exposed to alloxan also show significantly increased mitochondrial membrane potential. Apparently, alloxan causes severe oxidative and cytotoxic stress to islets that is likely to compromise their insulin releasing capacity [25].Overproduction of ROS or exhaustion of antioxidants may cause oxidative stress which is a major factor of defective insulin secretion and increases apoptosis of pancreas [26]. Moreover, ROS produced by β-cell in response to metabolic stress affect mitochondrial structure and function and lead to β-cell failure. Specifically, ROS oxidize mitochondrial membrane phospholipids such as cardiolipin, which impairs membrane integrity and leads to cytochrome c release and apoptosis. In addition, ROS activate UCP2 via peroxidation of the mitochondrial membrane phospholipids, which results in proton leak leading to reduced ATP synthesis and content in β-cells [27].However, continuous treatment of diabetic animals with CME for 30 days caused a significant reduction in FBG level. This antihyperglycemic action may be due to sulphated polysaccharides from marine algae that are known to be important free-radical scavengers and antioxidants for the prevention of pancreatic oxidative damage, which is an important contributor in DM [28a]. The observed depletion of liver glycogen stores in diabetic rats is consistent with an earlier study [29] indicating that it could be due to the loss of glycogen-synthase activating system and/or increased activity of glycogen phosphorylase [30] since alloxan causes selective destruction of β-cells of the islets of langerhans resulting in a marked decrease of insulin levels, it is rational to believe that the activity of hepatic glycogen phosphorylase and hepatic glucose-6- phosphatase of diabetic animals will increased, as synthesis of glycogen depends on insulin for the influx of glucose. However, previous studies show that the reduced activity of glycogen phosphorylase and hepatic glucose-6- phosphatase hepatic by insulin treatment [31].Administration of CME for 30 days also showed a trend towards a significant increase in glycogen content when compared to diabetic rats, thus confirming its insulin potentiating effect to a certain extent.The antihyperglycemic efficacy of this herbal drug can be correlated with the findings of [32]. Similarly, alloxan caused body weight loss was also regained to its above-initial values by CME treatment, which reflects an improved health of CME treated animals. The histological analysis of the pancreas further substantiates a conclusion that CME has a protective effect on pancreatic tissueThe characteristic features of diabetic dyslipidemia are a high plasma triglyceride concentration, low HDL-cholesterol concentration and increased concentration of small dense LDL-cholesterol particles. Faulty glucose utilization causes hyperglycemia and mobilization of fatty acids from adipose tissue for energy purpose. The lipid changes associated with diabetes mellitus are attributed to increased flux of free fatty acids into the liver secondary to insulin deficiency/ resistance. This results in excess fatty acid accumulation in the liver, which is converted to triglycerides. The impaired ability of insulin to inhibit free fatty-acid release leads to elevated hepatic VLDL-cholesterol production. The increased VLDL-cholesterol and triglyceride levels decrease the level of HDL-cholesterol and increase the concentration of small dense LDL-cholesterol particles by activation of lipoprotein lipase and lecithin acyl-cholesterol transferase We found elevated levels of serum TC, TG, LDL cholesterol and decreased HDL cholesterol concentration in alloxan-induced diabetic rats [33a]. On other hand, Induction of diabetes by alloxan resulted in loss of body weight in the diabetic control rats that may due to catabolic effect on protein metabolism by retarding protein synthesis and stimulating protein degradation. However, treatment with CME normalized all the lipid profile parameters. This antihyperlipidemic attributed to sulphated poly saccharides , major constituent of extract as sulphated poly saccharides enhance the negative charges of cell surface so as to effect the aggradation of cholesterol in blood, thus decreasing the cholesterol in serum [34].Earlier it has been explored that oxidative stress forms the foundation for the induction of multiple cellular pathways that can ultimately lead to both the onset and subsequent complications of DM. Diabetics and experimental animal models exhibit high oxidative stress due to persistent and chronic hyperglycemia, which thereby depletes the activity of antioxidative defense system and thus promotes de novo free radicals generation [4b].GSH oxidation is a major contributor to cell apoptosis mediated by oxidants. Accumulated evidence from our laboratory has consistently shown that an early spike in GSSG formation, typically within minutes of oxidant exposure, preceded oxidant-induced activation of mitochondrial apoptotic signaling and cell apoptosis hours later [35].In our study, the level of hepatic and pancreatic GSH was reduced in diabetic rats, which is consistent with an earlier report [36].The decrease in tissue’s GSH content could be the result of decreased synthesis or increased degradation of GSH by oxidative stress that prevails duringdiabetes [37]. Furthermore, lipid peroxidation is one of the characteristic features of chronic diabetes [33b]. In the present study, along with decreased level of reduced GSH, a marked increase in the concentration of TBARS was also observed in liver and pancreas of diabetic mice; however, treatment of these animals with CME decreased the elevated level of TBARS. Simultaneously, reduced GSH content was also increased significantly which indicates that the CME can either increase the biosynthesis of GSH and/or reduce the oxidative stress that ultimately reduces the degradation of GSH. Similar results were also witnessed in an earlier study [38].Catalase is a common enzyme found in nearly all living organisms. Its functions include catalyzing the decomposition of hydrogen peroxide to water and oxygen. One of the most commonly used ROS for neuronal oxidative stress preconditioning is H2O2. It is formed by the dismutation of superoxide (O2) spontaneously or enzymatically in a reaction that is catalyzed by superoxide dismutase (SOD). H2O2 is less reactive compared to other ROS, easily crosses membranes, and diffuses from its original site of production.Many more studies documented a pivotal role for H2O2 in the development of vascular dysfunction in pathological conditions, such as atherosclerosis, hypertension, and diabetes mellitus. It has also been speculated that, in the vessel wall, H2O2-mediated mechanisms may compensate for the loss of NO-mediated dilation during the development of various diseases [39].To prevent damage, it must be quickly converted into other, less dangerous substances. To this end, catalase is frequently used by cells to rapidly catalyze the decomposition of hydrogen peroxide into less reactive gaseous oxygen and water molecules. Superoxide dismutases are a class of enzymes that catalyze the dismutation of superoxide into oxygen and hydrogen peroxide. As such, they are an important antioxidant defense in nearly all cells exposed to oxygen.However, CME therapy increased the activity of CAT and GSH-Px in pancreatic tissues of treated diabetic animals [28b]. Reductone-associated sulphonic groups of polysaccharides can act as electron donors and can react with free radicals to convert these polysaccharides to more stable products and thereby terminate radical chain reactions [40].The increased activity of CAT, SOD and GSH-Px suggest a compensatory response to oxidative stress as it reduces the endogenous H2O2 produced thus diminishing the toxic effects due to this radical or other free radicals derived from secondary reactions [41, 42].Moreover, the observed antioxidative effect of CME may due to synergistic action of various compounds present in the extract in contrast to the purified compound of the drug.However, the level of Hepatic TBARS, Hepatic GSH and activity of antioxidant enzymes in hepatic tissue of diabetic rats was not improved by CME treatment. This perhaps is due to different metabolic actions of the tissues and their different responses to oxidative stress or the hepatotoxic nature of the extract.The histological observation of the hepatic tissues further substantiates the claim that CME has a toxic nature on hepatic tissue.Results of this study do not allow definite conclusion to be drawn on the mechanism of action of CME in the experimental animal paradigms used. However, a number of investigators have shown that a host of various plant secondary metabolites possess hypoglycemic, hypotensive, anti-inflammatory and other pharmacological properties [7b]. As mentioned earlier, CME is also known to contain various secondary metabolites. Therefore, it is not unreasonable to speculate that some of these chemical compounds are presumably responsible for imparting the antihyperglycemic, antihyperlipidemic and antioxidative properties to CME.
5. Conclusions
- From the results obtained, it can be concluded that CME possess significant antihyperglycemic, antihyperlipidemic and pancreatic antioxidative properties. Hence, apart from controlling hyperglycemia it would also be beneficial in the alleviation of associated diabetic complications including the prevention of the development of atherosclerosis and other coronary artery diseases. However, further studies are needed to investigate and elucidate the possible mechanism of action of the active ingredients, establish complete safety profiles and evaluate the potential value of CME for the management of diabetes and hyperlipidemia in the clinic.Moreover, additional parameters such as assay of fructosamine, HbA1c, C-peptide etc should also be studied. This may prove helpful for developing new drugs from this plant for managing diabetes and associated complications.
ACKNOWLEDGEMENTS
- Authors gratefully acknowledge Vice Chancellor, El Minia, for the financial support through the Department of Agriculture chemistry.
References
| [1] | Park MK, Jung U, Roh C (2011): Fucoidan from marine brown algae inhibits lipid accumulation. Drugs, 9(8):1359-67. |
| [2] | Matsui T, Tanaka T, Tamura S, Toshima A, Miyata Y, Tanaka K (2007): Alphaglucosidase inhibitory profile of catechins and thioflavins. Journal of Agricultural and Food Chemistry, 55, 99–105. |
| [3] | Maiese K, Chong ZZ, Shang YC (2007): Mechanistic insights into diabetes mellitus and oxidative stress. Curr Med Chem; 14(16):1729-38. |
| [4] | Mellor KM, Ritchie RH, Delbridge LM (2010): Reactive oxygen species and insulin-resistant cardiomyopathy. Clin Exp Pharmacol Physiol, 37(2):222-8. |
| [5] | Matough FA, Budin SB, Hamid ZA, Alwahaibi N , Mohamed J(2012): The role of oxidative stress and antioxidants in diabetic complications. Sultan Qaboos Univ Med J; 12(1):5-18. |
| [6] | Amin KA , Nagy MA (2009): Effect of carnitine and herbal mixture exract on obesity inducedby high fat diet in rats. Diabetol Metab Syndr. 16; 1(1):17. |
| [7] | Wang H, Chiu LCM, Ooi VEC, Ang Jr PO (2010): A potent antitumor polysaccharide from the edible brown seaweed Hydroclathrus clathratus, Bot. 53 265–274. |
| [8] | Sadati N, Khanavi M Mahrokh A, Nabavi SMB , Sohrabipour J, Hadjiakhoondi A (2011): Comparison of Antioxidant Activity and Total Phenolic Contents of some Persian Gulf Marine Algae. Journal of Medicinal Plants, Volume 10, No. 37. |
| [9] | Jayasooriya RG, Choi YH, Moon SK, Kim WJ, Kim GY (2012): Methanol extract of Hydroclathrus clathratus suppresses matrix metalloproteinase-9 in T24 bladder carcinoma cells by suppressing the NF-κB and MAPK pathways. Oncol Rep; 27(2):541-6. |
| [10] | Viswanathaswamy AH, Koti BC, Gore A, Thippeswamy AH and Kulkarni RV (2011): Antihyperglycemic and antihyperlipidemic activity of plectranthus amboinicus on normal and alloxan-induced diabetic rats. Indian J Pharm Sci. ;73(2):139-45. |
| [11] | Trinder, PPlasma glucose was measured by standard methods. (1969), Annals of Clinical Biochemistry, 6, 24–27. |
| [12] | Marschner I, Bottermann P, Erhardt F, Linke R, Löffler G, Maier V, Schwandt P, VogtW , Scriba PC (1974) : "Group experiments on the radioimmunological insulin determination" Horm Metab Res. l;6(4):293-6. |
| [13] | Kakkar P, Das B, Viswanathan PN (1984)A modified spectrophotometric assay of superoxide dismutase. Ind J Biochem Biophys; 21: 130-32. |
| [14] | Lowry OH, Rosebrough, NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem; 193: 265-75. |
| [15] | Sinha AK (1972) Colorimetric assay of catalase. Anal Biochem; 47: 389-94. |
| [16] | Rotruck JT, Pope AL, Ganther HF, Swanson AB (1973). Selenium: Biochemical role as a component of glutathione peroxidase. Science; 179: 588-90. |
| [17] | Begum N, Sam GPK, Shanmugasundaram KR (1978) Serum enzymes I human and experimental diabetes mellitus. Indian J Med Res.; 68:774-784. |
| [18] | Ohkawa H, Ohishi N, Yagi K (1979): Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem; 95: 351–58. |
| [19] | Stalmans W, Hers HG (1975) The stimulation of liver glycogen phosphorylase b by AMP, fluoride and sulfate. A technical note of the specific determination of the a and b forms of liver glycogen phosphorylase Eur j Biochem; 54:341-350. |
| [20] | Donath MY, Halban PA (2004): Decreased beta-cell mass in diabetes: significance, mechanisms and therapeutic implications. Diabetologia47:581 –589. |
| [21] | Szkudelski T (2001): The mechanism of alloxan and streptozotocin action in β-cells of the rat pancreas.Physiol Res.; 50:536–46. |
| [22] | Johansen JS, Harris AK, Rychly DJ, Ergul A (2005): Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical practice. Cardiovasc Diabetol; 4:1–11. |
| [23] | Ceriello A (2000): Oxidative stress and glycemic regulation. 49:27-9. |
| [24] | Lebedev VP, Bilichenko SV, Ordyan NE, Pivina SG, Nechiporenko SP, Puzyrev AA, Mikheeva EA, Kubacheva KK (2007): Transcranial electrostimulation activates reparative regeneration and the insulin-producing function of pancreatic B-cells in alloxan diabetes in rats. Neurosci Behav Physiol. 37(4):341-7. |
| [25] | Rabbani SI, Devi K, Khanam S (2010): Role of Pioglitazone with Metformin or Glimepiride on Oxidative Stress-induced Nuclear Damage and Reproductive Toxicity in Diabetic Rats. Malays J Med Sci. 17(1):3-11. |
| [26] | Lee E, Ryu GR, Ko SH, Ahn YB, Yoon KH, Ha H , Song KH (2011): Antioxidant treatment may protect pancreatic beta cells through the attenuation of islet fibrosis in an animal model of type 2 diabetes. Biochem Biophys Res Commun. 22; 414(2):397-402. |
| [27] | DU SC, Ge QM, Lin N, Dong Y , Su Q (2012): Ros-mediated lipopolysaccharide-induced apoptosis in ins-1 cells by modulation of bcl-2 and bax. Cell Mol Biol (Noisy-le-grand). 23; 58. |
| [28] | Ma ZA (2012): The Role of Peroxidation of Mitochondrial Membrane Phospholipids in Pancreatic β -Cell Failure. Curr Diabetes Rev. 1; 8(1):69-75. |
| [29] | Huang ZX, Mei XT,Xu DH, Xu SB, Lv JY (2005): Protective effects of polysacchride of Spirulina platensis and Sargassum thunbeergii on vascular of alloxan induced diabetic rats. Zhongguo Zhong Yao Za Zhi. 30(3):211-5. |
| [30] | Hamden K, Carreau S, Jamoussi K, Miladi S, Lajmi S, Aloulou D, Ayadi F, Elfeki A (2009): 1Alpha, 25 dihydroxyvitamin D3: therapeutic and preventive affects against oxidative stress, hepatic, pancreatic and renal injury in alloxan-induced diabetes in rats. J Nutr Sci Vitaminol (Tokyo). 55(3):215-22. |
| [31] | Abdel-Moneim A, El-Feki M, Salh E (1997) Effect of Nigella sativa, fish oil and glicazide on alloxan diabetic rats. 1- Biochemical and Histopathological studies. J Egyp Ger Soc Zoo; 23: 237-65. |
| [32] | El-Shenawy NS, Abdel-Nabi IM (2006) Hypoglycemic effect of Cleome droserifolia ethanolic leaf extract in experimental diabetes, and on non-enzymatic antioxidant, glycogen, thyroid hormone and insulin levels. Diabetologia Croatica; 35: 15-22. |
| [33] | Liu H, Gu L (2012): Phlorotannins from brown algae (Fucus vesiculosus) inhibited the formation of advanced glycation endproducts by scavenging reactive carbonyls. . J Agric Food Chem. 8; 60(5): 1326-34. |
| [34] | Yadav UC, Moorthy K, Baquer NZ (2005): Combined treatment of sodium orthovanadate and Momordica charantia fruit extract prevents alterations in lipid profile and lipogenic enzymes in alloxan diabetic rats. Mol Cell Biochem. 268(1-2):111-20. |
| [35] | Kim SK, Li YX (2011): Medicinal benefits of sulfated polysaccharides from sea vegetables. Adv Food Nutr Res. 64:391-402. |
| [36] | Pias EK, Ekshyyan OY, Rhoads CA, Fuseler J, Harrison L , Aw TY (2003): Differential effects of superoxide dismutase isoform expression on hydroperoxide-induced apoptosis in PC-12 cells. J Biol Chem., 278:13294–13301. |
| [37] | Brito VB, da Rocha JB, Puntel GO, da Luz SC, Barbosa NB, de Carvalho NR, Folmer V (2011): Inhibition of δ-aminolevulinate dehydratase is not closely related to the development of hyperglycemia in alloxan-induced diabetic mice. Exp Toxicol Pathol. 63(5):443-51. |
| [38] | Soto C, Pérez J, García V, Uría E, Vadillo M , Raya L (2010): Effect of silymarin on kidneys of rats suffering from alloxan-induced diabetes mellitus. Phytomedicine. 1; 17(14):1090-4. |
| [39] | Hayashi S, Itoh A Isoda K , Kondoh M , Kawase M , Yagi K (2008): Fucoidan partly prevents CCl4-induced liver fibrosis. Eur. J. Pharmacol , 580, 380-384. |
| [40] | Kobayashi T, Kamata K (2002): Modulation by hydrogen peroxide of noradrenaline-induced contraction in aorta from streptozotocin-induced diabetic rat. Eur J Pharmacol 441: 83–89. |
| [41] | Hlawaty H , Suffee N , Sutton A , Oudar O , Haddad O , Ollivier V, Laguillier-Morizot C , Gattegno L , Letourneur D , Charnaux N(2011): Low molecular weight fucoidan prevents intimal hyperplasia in rat injured thoracic aorta through the modulation of matrix metalloproteinase-2 expression. Biochem. Pharmacol , 81, 233–243. |
| [42] | Aisa Y, Miyakawa Y, Nakazato T, Shibata H, Saito K , Ikeda Y (2004): Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. American Journal of Hematology, 78, 7–14. |
| [43] | Hu JF, Geng MY, Zhang JT, Jiang HD (2001): An in vitro study of the structure-activity relations of sulfated polysaccharides from brown algae to its antioxidant effect. J Asian Nat Prod Reports 3:353-358. |